

PLP/DM20 ratio is regulated by hnRNPH and F and a novel G-rich enhancer in oligodendrocytes
- PMID: 17567613
- PMCID: PMC1919487
- DOI: 10.1093/nar/gkm387
PLP/DM20 ratio is regulated by hnRNPH and F and a novel G-rich enhancer in oligodendrocytes
Abstract
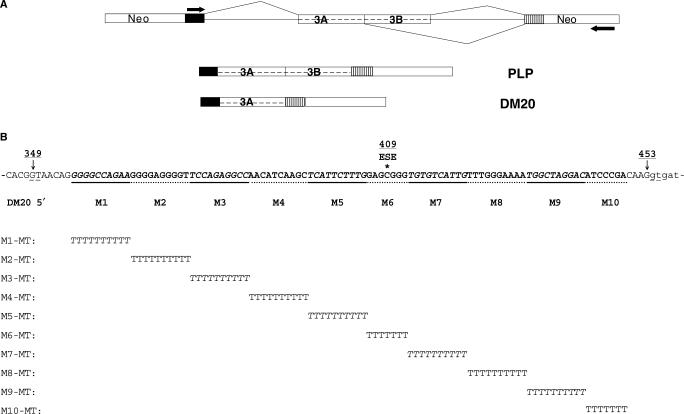
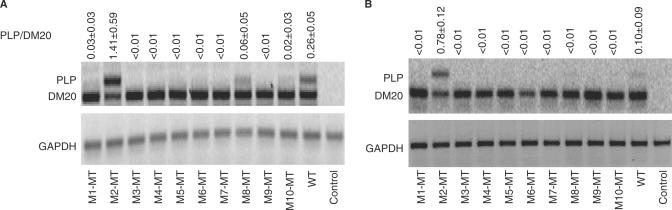
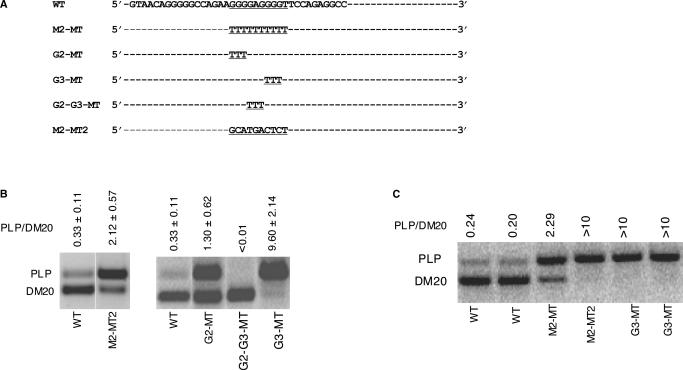
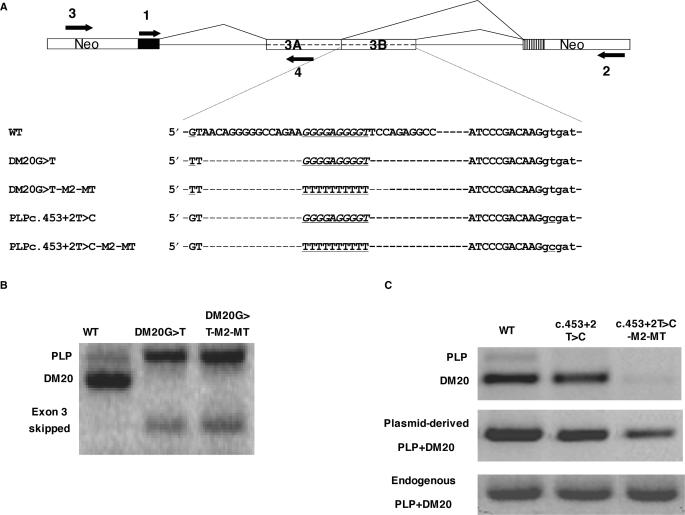
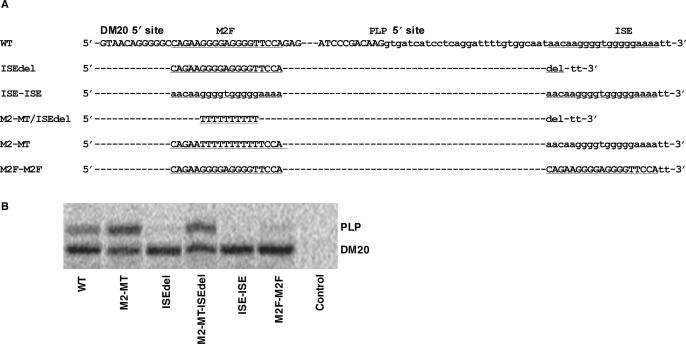
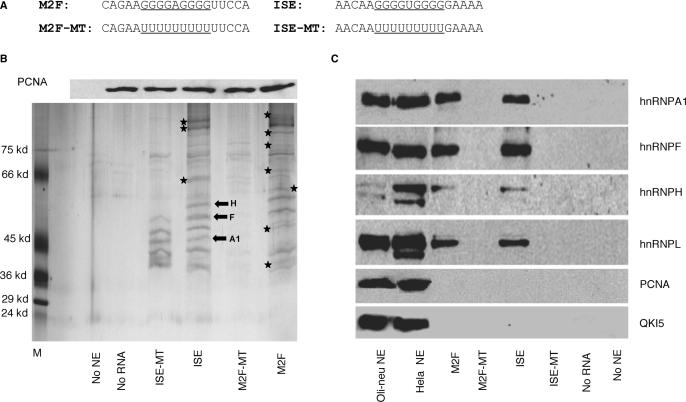
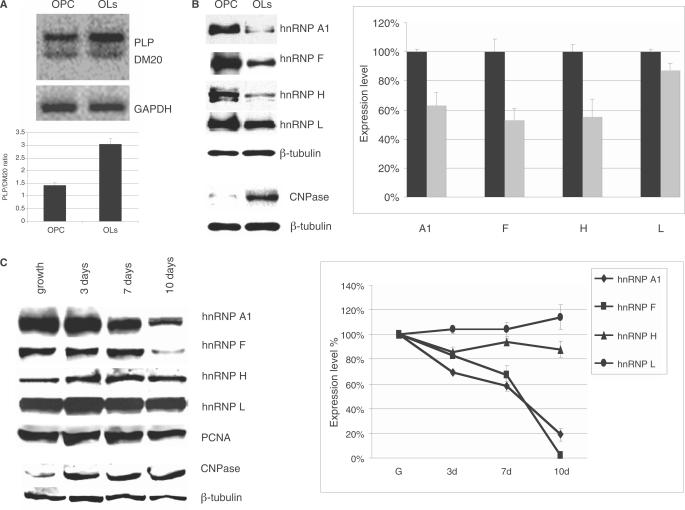
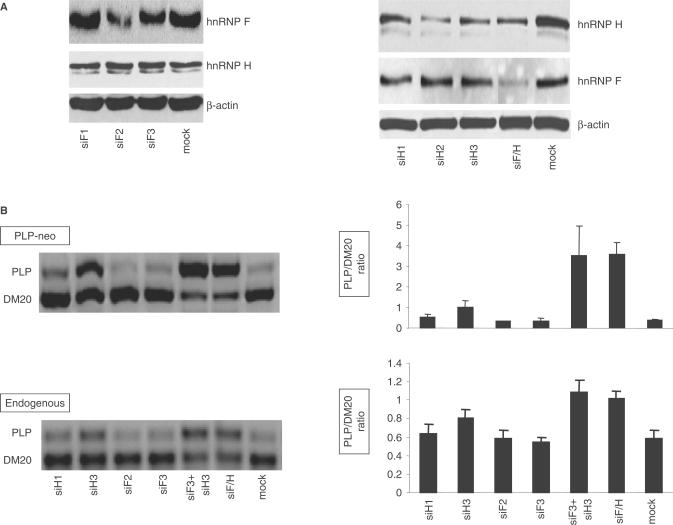
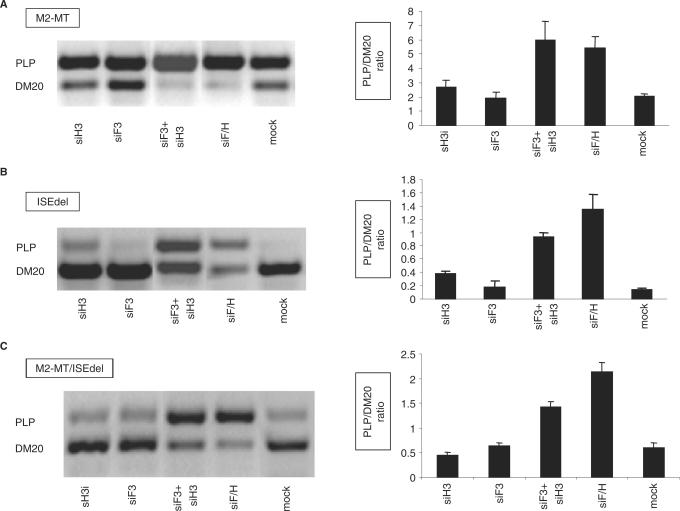
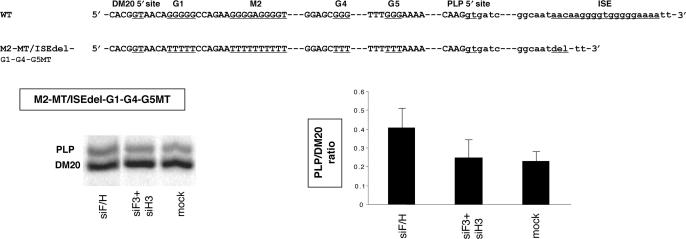
Alternative splicing of competing 5' splice sites is regulated by enhancers and silencers in the spliced exon. We have characterized sequences and splicing factors that regulate alternative splicing of PLP and DM20, myelin proteins produced by oligodendrocytes (OLs) by selection of 5' splice sites in exon 3. We identify a G-rich enhancer (M2) of DM20 5' splice site in exon 3B and show that individual G triplets forming M2 are functionally distinct and the distal group plays a dominant role. G-rich M2 and a G-rich splicing enhancer (ISE) in intron 3 share similarities in function and protein binding. The G-rich sequences are necessary for binding of hnRNPs to both enhancers. Reduction in hnRNPH and F expression in differentiated OLs correlates temporally with increased PLP/DM20 ratio. Knock down of hnRNPH increased PLP/DM20 ratio, while hnRNPF did not. Silencing hnRNPH and F increased the PLP/DM20 ratio more than hnRNPH alone, demonstrating a novel synergistic effect. Mutation of M2, but not ISE reduced the synergistic effect. Replacement of M2 and all G runs in exon 3B abolished it almost completely. We conclude that developmental changes in hnRNPH/F associated with OLs differentiation synergistically regulate PLP alternative splicing and a G-rich enhancer participates in the regulation.
Figures
References
-
- Modrek B, Lee CJ. Alternative splicing in the human, mouse and rat genomes is associated with an increased frequency of exon creation and/or loss. Nat. Genet. 2003;34:177–180. - PubMed
-
- Sugnet CW, Kent WJ, Ares M, Jr, Haussler D. Transcriptome and genome conservation of alternative splicing events in humans and mice. Pac. Symp. Biocomput. 2004:66–77. - PubMed
-
- Matlin AJ, Clark F, Smith CW. Understanding alternative splicing: towards a cellular code. Nat. Rev. Mol. Cell Biol. 2005;6:386–398. - PubMed
-
- Black DL, Grabowski PJ. Alternative pre-mRNA splicing and neuronal function. Prog. Mol. Subcell. Biol. 2003;31:187–216. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
