

Transcription factor binding and modified histones in human bidirectional promoters
- PMID: 17568000
- PMCID: PMC1891341
- DOI: 10.1101/gr.5623407
Transcription factor binding and modified histones in human bidirectional promoters
Abstract
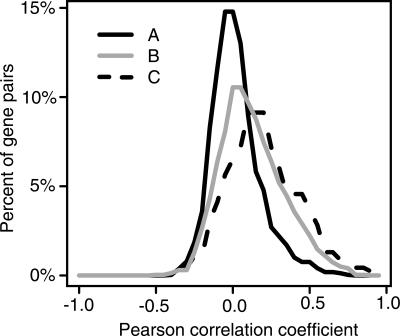
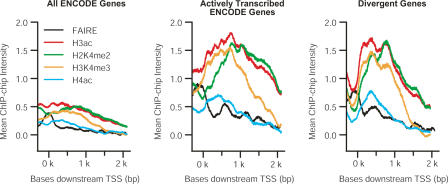
Bidirectional promoters have received considerable attention because of their ability to regulate two downstream genes (divergent genes). They are also highly abundant, directing the transcription of approximately 11% of genes in the human genome. We categorized the presence of DNA sequence motifs, binding of transcription factors, and modified histones as overrepresented, shared, or underrepresented in bidirectional promoters with respect to unidirectional promoters. We found that a small set of motifs, including GABPA, MYC, E2F1, E2F4, NRF-1, CCAAT, YY1, and ACTACAnnTCC are overrepresented in bidirectional promoters, while the majority (73%) of known vertebrate motifs are underrepresented. We performed chromatin-immunoprecipitation (ChIP), followed by quantitative PCR for GABPA, on 118 regions in the human genome and showed that it binds to bidirectional promoters more frequently than unidirectional promoters, and its position-specific scoring matrix is highly predictive of binding. Signatures of active transcription, such as occupancy of RNA polymerase II and the modified histones H3K4me2, H3K4me3, and H3ac, are overrepresented in regions around bidirectional promoters, suggesting that a higher fraction of divergent genes are transcribed in a given cell than the fraction of other genes. Accordingly, analysis of whole-genome microarray data indicates that 68% of divergent genes are transcribed compared with 44% of all human genes. By combining the analysis of publicly available ENCODE data and a detailed study of GABPA, we survey bidirectional promoters with breadth and depth, leading to biological insights concerning their motif composition and bidirectional regulatory mode.
Figures






References
-
- Adachi N., Lieber M.R., Lieber M.R. Bidirectional gene organization: A common architectural feature of the human genome. Cell. 2002;109:807–809. - PubMed
-
- Bernstein B.E., Kamal M., Lindblad-Toh K., Bekiranov S., Bailey D.K., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Kamal M., Lindblad-Toh K., Bekiranov S., Bailey D.K., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Lindblad-Toh K., Bekiranov S., Bailey D.K., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Bekiranov S., Bailey D.K., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Bailey D.K., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Huebert D.J., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., McMahon S., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Karlsson E.K., Kulbokas E.J., 3rd, Gingeras T.R., Kulbokas E.J., 3rd, Gingeras T.R., Gingeras T.R., et al. Genomic maps and comparative analysis of histone modifications in human and mouse. Cell. 2005;120:169–181. - PubMed
-
- Bieda M., Xu X., Singer M.A., Green R., Farnham P.J., Xu X., Singer M.A., Green R., Farnham P.J., Singer M.A., Green R., Farnham P.J., Green R., Farnham P.J., Farnham P.J. Unbiased location analysis of E2F1-binding sites suggests a widespread role for E2F1 in the human genome. Genome Res. 2006;16:595–605. - PMC - PubMed
-
- Boyle E.I., Weng S., Gollub J., Jin H., Botstein D., Cherry J.M., Sherlock G., Weng S., Gollub J., Jin H., Botstein D., Cherry J.M., Sherlock G., Gollub J., Jin H., Botstein D., Cherry J.M., Sherlock G., Jin H., Botstein D., Cherry J.M., Sherlock G., Botstein D., Cherry J.M., Sherlock G., Cherry J.M., Sherlock G., Sherlock G. GO::TermFinder–open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics. 2004;20:3710–3715. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials