

Interaction of HapX with the CCAAT-binding complex--a novel mechanism of gene regulation by iron
- PMID: 17568774
- PMCID: PMC1914100
- DOI: 10.1038/sj.emboj.7601752
Interaction of HapX with the CCAAT-binding complex--a novel mechanism of gene regulation by iron
Abstract
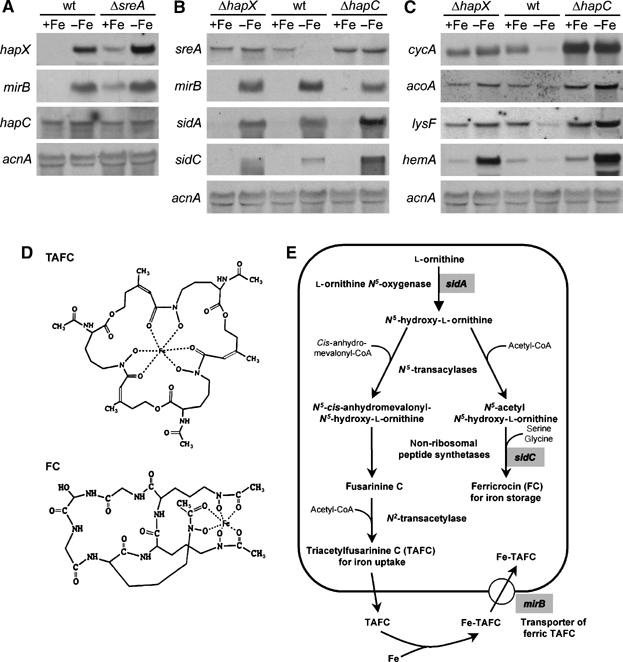
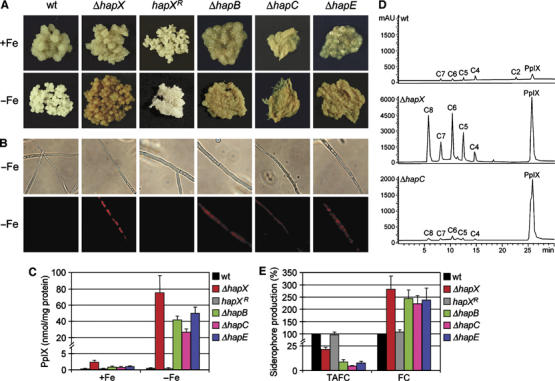
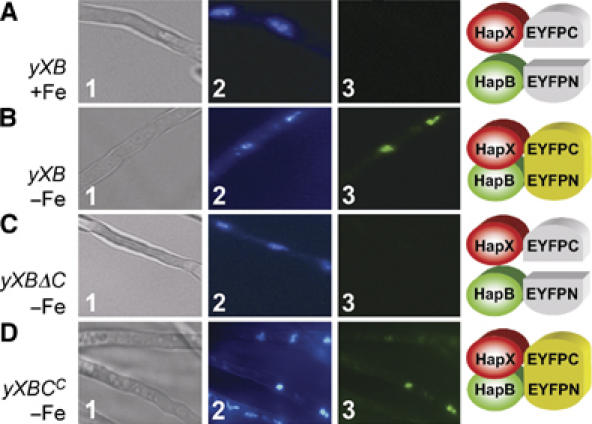
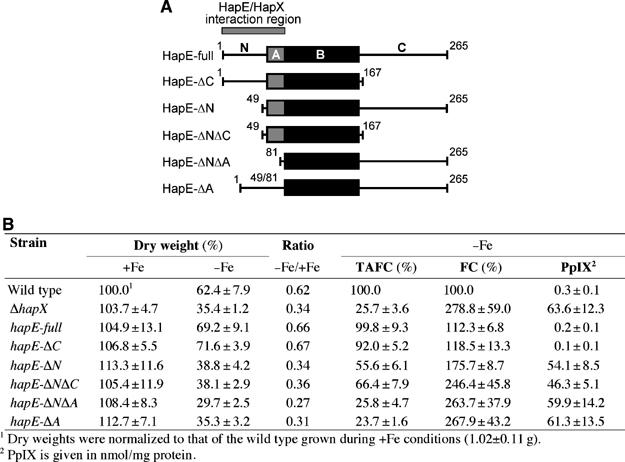
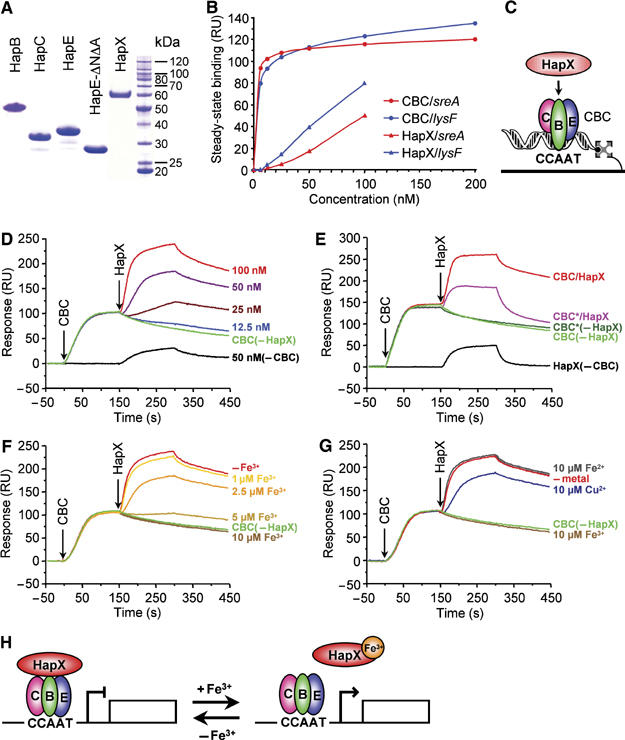
Iron homeostasis requires subtle control systems, as iron is both essential and toxic. In Aspergillus nidulans, iron represses iron acquisition via the GATA factor SreA, and induces iron-dependent pathways at the transcriptional level, by a so far unknown mechanism. Here, we demonstrate that iron-dependent pathways (e.g., heme biosynthesis) are repressed during iron-depleted conditions by physical interaction of HapX with the CCAAT-binding core complex (CBC). Proteome analysis identified putative HapX targets. Mutual transcriptional control between hapX and sreA and synthetic lethality resulting from deletion of both regulatory genes indicate a tight interplay of these control systems. Expression of genes encoding CBC subunits was not influenced by iron availability, and their deletion was deleterious during iron-depleted and iron-replete conditions. Expression of hapX was repressed by iron and its deletion was deleterious during iron-depleted conditions only. These data indicate that the CBC has a general role and that HapX function is confined to iron-depleted conditions. Remarkably, CBC-mediated regulation has an inverse impact on the expression of the same gene set in A. nidulans, compared with Saccharomyces cerevisae.
Figures
References
-
- Bonkovsky HL, Wood SG, Howell SK, Sinclair PR, Lincoln B, Healey JF, Sinclair JF (1986) High-performance liquid chromatographic separation and quantitation of tetrapyrroles from biological materials. Anal Biochem 155: 56–64 - PubMed
-
- Bourgarel D, Nguyen CC, Bolotin-Fukuhara M (1999) HAP4, the glucose-repressed regulated subunit of the HAP transcriptional complex involved in the fermentation-respiration shift, has a functional homologue in the respiratory yeast Kluyveromyces lactis. Mol Microbiol 31: 1205–1215 - PubMed
-
- Bradshaw RE, Dixon SW, Raitt DC, Pillar TM (1993) Isolation and nucleotide sequence of the 5-aminolevulinate synthase gene from Aspergillus nidulans. Curr Genet 23: 501–507 - PubMed
-
- Brakhage AA, Andrianopoulos A, Kato M, Steidl S, Davis MA, Tsukagoshi N, Hynes MJ (1999) HAP-like CCAAT-binding complexes in filamentous fungi: implications for biotechnology. Fungal Genet Biol 27: 243–252 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
