

Sex-dependent metabolic, neuroendocrine, and cognitive responses to dietary energy restriction and excess
- PMID: 17569758
- PMCID: PMC2622430
- DOI: 10.1210/en.2007-0161
Sex-dependent metabolic, neuroendocrine, and cognitive responses to dietary energy restriction and excess
Abstract
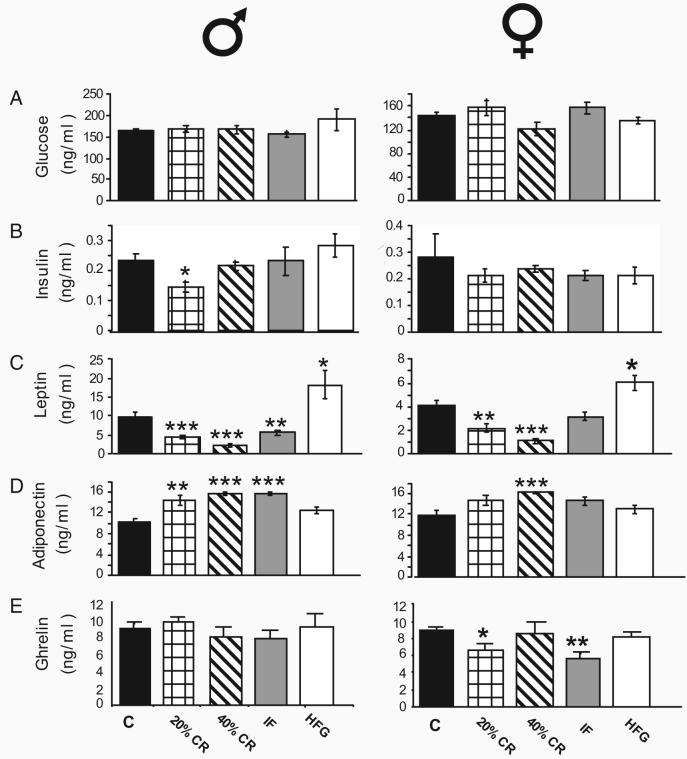
Females and males typically play different roles in survival of the species and would be expected to respond differently to food scarcity or excess. To elucidate the physiological basis of sex differences in responses to energy intake, we maintained groups of male and female rats for 6 months on diets with usual, reduced [20% and 40% caloric restriction (CR), and intermittent fasting (IF)], or elevated (high-fat/high-glucose) energy levels and measured multiple physiological variables related to reproduction, energy metabolism, and behavior. In response to 40% CR, females became emaciated, ceased cycling, underwent endocrine masculinization, exhibited a heightened stress response, increased their spontaneous activity, improved their learning and memory, and maintained elevated levels of circulating brain-derived neurotrophic factor. In contrast, males on 40% CR maintained a higher body weight than the 40% CR females and did not change their activity levels as significantly as the 40% CR females. Additionally, there was no significant change in the cognitive ability of the males on the 40% CR diet. Males and females exhibited similar responses of circulating lipids (cholesterols/triglycerides) and energy-regulating hormones (insulin, leptin, adiponectin, ghrelin) to energy restriction, with the changes being quantitatively greater in males. The high-fat/high-glucose diet had no significant effects on most variables measured but adversely affected the reproductive cycle in females. Heightened cognition and motor activity, combined with reproductive shutdown, in females may maximize the probability of their survival during periods of energy scarcity and may be an evolutionary basis for the vulnerability of women to anorexia nervosa.
Figures








References
-
- Levine A, Billington C. Why do we eat? A neural systems approach. Annu Rev Nutr. 1997;17:597–619. - PubMed
-
- Badman M, Flier J. The gut and energy balance: visceral allies in the obesity wars. Science. 2005;307:1909–1914. - PubMed
-
- Volkow N, Wise R. How can drug addiction help us understand obesity? Nat Neurosci. 2005;8:555–560. - PubMed
-
- Haslam D, James W. Obesity. Lancet. 2005;366:1197–1209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
