

Hematopoietic stem-cell gene therapy of hemophilia A incorporating a porcine factor VIII transgene and nonmyeloablative conditioning regimens
- PMID: 17569821
- PMCID: PMC2018667
- DOI: 10.1182/blood-2007-04-082602
Hematopoietic stem-cell gene therapy of hemophilia A incorporating a porcine factor VIII transgene and nonmyeloablative conditioning regimens
Abstract
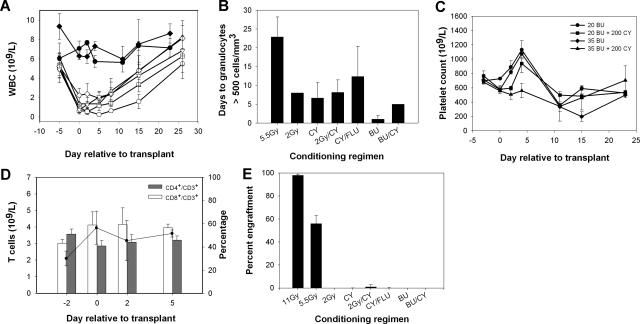
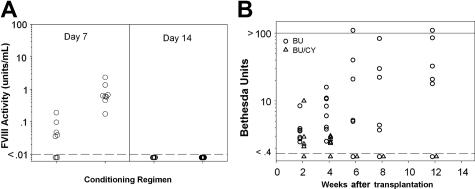
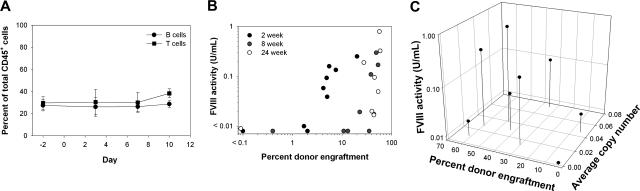
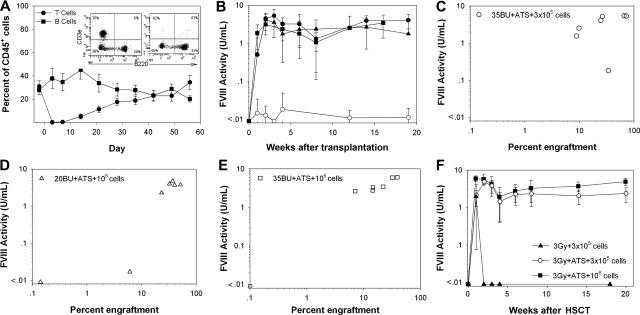
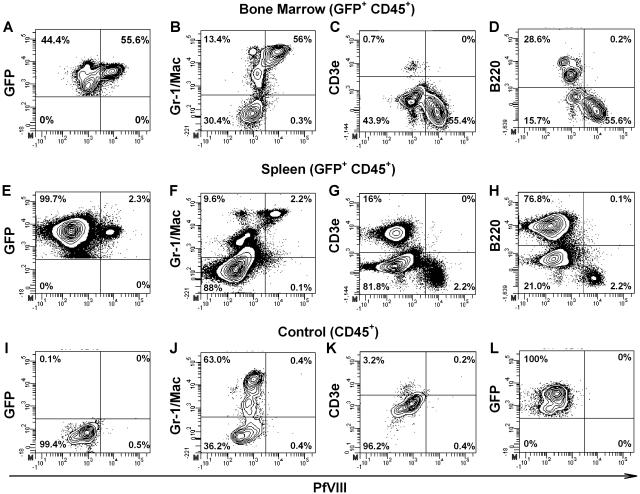
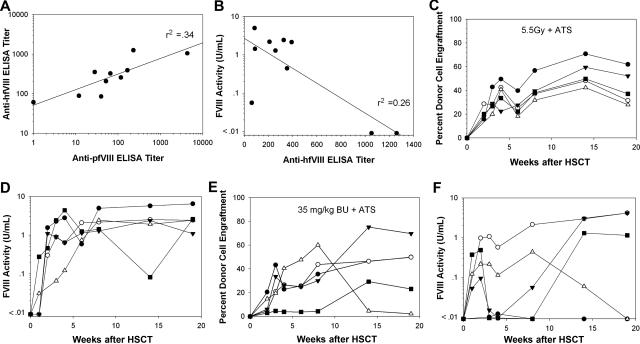
Insufficient expression of factor VIII (fVIII) is a major hurdle in the development of successful nucleic acid treatments for hemophilia. However, we recently showed that under myeloablative and reduced-intensity total body irradiation (TBI) conditioning, transplantation of hematopoietic stem cells (HSCs) transduced with recombinant retroviruses containing B domain-deleted porcine fVIII (BDDpfVIII) sequences provides curative fVIII levels in a hemophilia A mouse model. In the current study, we tested BDDpfVIII activity after nonmyeloablative conditioning with busulfan, cyclophosphamide, or fludarabine and immunosuppressive agents CTLA4-Ig + anti-CD40L or anti-(murine)thymocyte serum (ATS). ATS is similar in action to anti-(human)thymocyte globulin (ATG), which is used clinically with busulfan in bone marrow transplantations to increase donor cell engraftment. Mice conditioned with busulfan + ATS and that received a transplant of BDDpfVIII-transduced stem-cell antigen 1-positive cells exhibited moderate levels of donor cell chimerism (between 20% and 60%) and achieved sustained fVIII levels more than 1 U/mL. Similar results were observed in mice preimmunized with human fVIII and conditioned with 5 Gy TBI + ATS or busulfan + ATS. These data demonstrate that it is possible to achieve sufficient fVIII expression after transplantation of BDDpfVIII-transduced HSCs following low-toxicity pretransplantation conditioning with targeted immunosuppression, potentially even in the context of preexisting inhibitors.
Figures
References
-
- Lollar P. Pathogenic antibodies to coagulation factors. Part one: factor VIII and factor IX. J Thromb Haemost. 2004;2:1082–1095. - PubMed
-
- High K. Gene transfer for hemophilia: can therapeutic efficacy in large animals be safely translated to patients? J Thromb Haemost. 2005;3:1682–1691. - PubMed
-
- Doering CB, Healey JF, Parker ET, Barrow RT, Lollar P. Identification of porcine coagulation factor VIII domains responsible for high level expression via enhanced secretion. J Biol Chem. 2004;279:6546–6552. - PubMed
-
- Gangadharan B, Parker ET, Ide LM, Spencer HT, Doering CB. High-level expression of porcine factor VIII from genetically modified bone marrow-derived stem cells. Blood. 2006;107:3859–3864. - PubMed
-
- Wognum AW, Eaves AC, Thomas TE. Identification and isolation of hematopoietic stem cells. Arch Med Res. 2003;34:461–475. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
