

Interactions of Plasmodium falciparum erythrocyte membrane protein 3 with the red blood cell membrane skeleton
- PMID: 17570341
- PMCID: PMC4768760
- DOI: 10.1016/j.bbamem.2007.04.027
Interactions of Plasmodium falciparum erythrocyte membrane protein 3 with the red blood cell membrane skeleton
Abstract
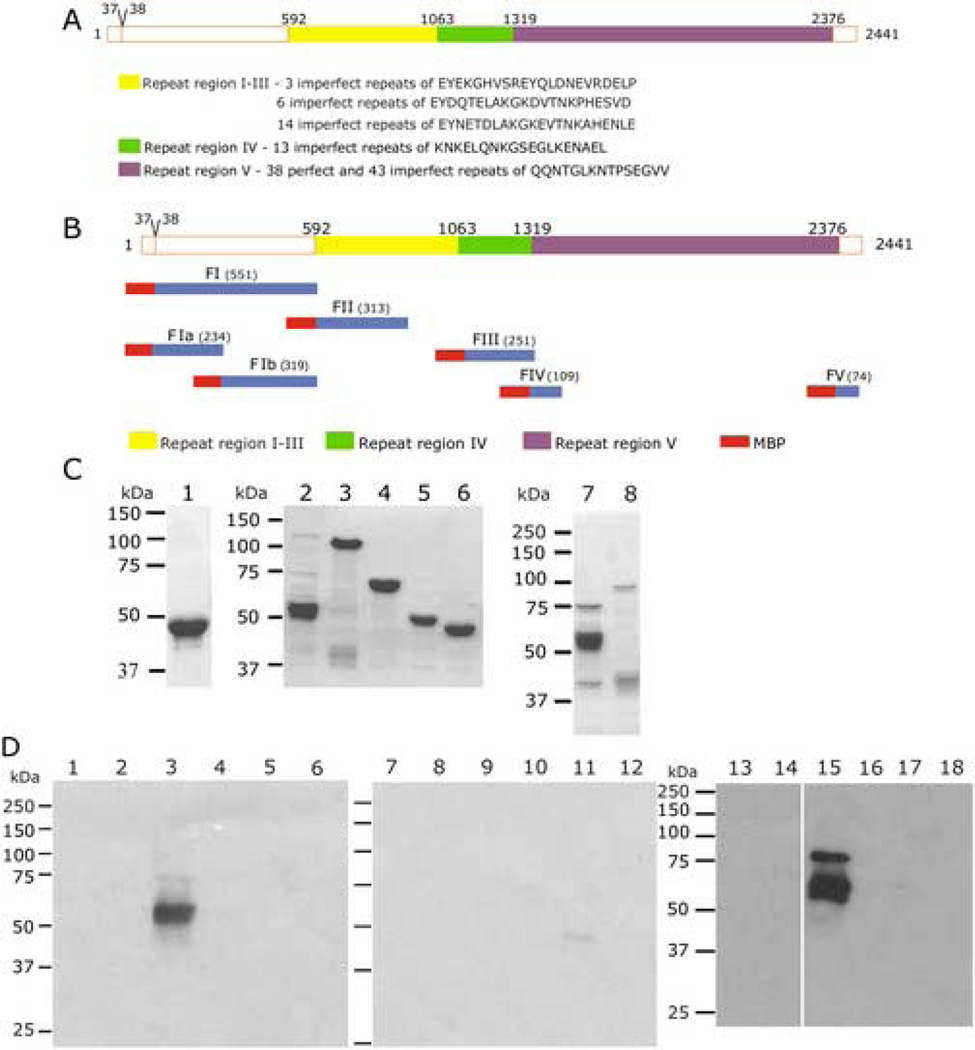
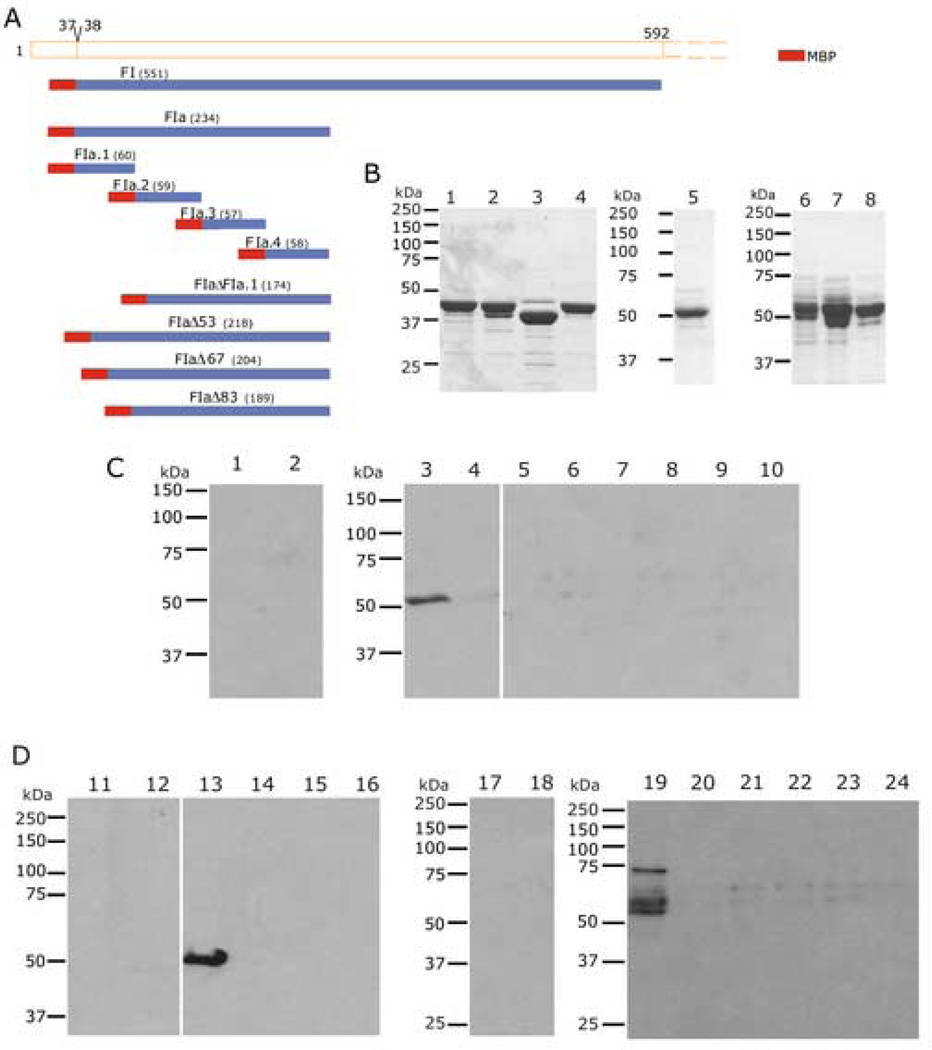
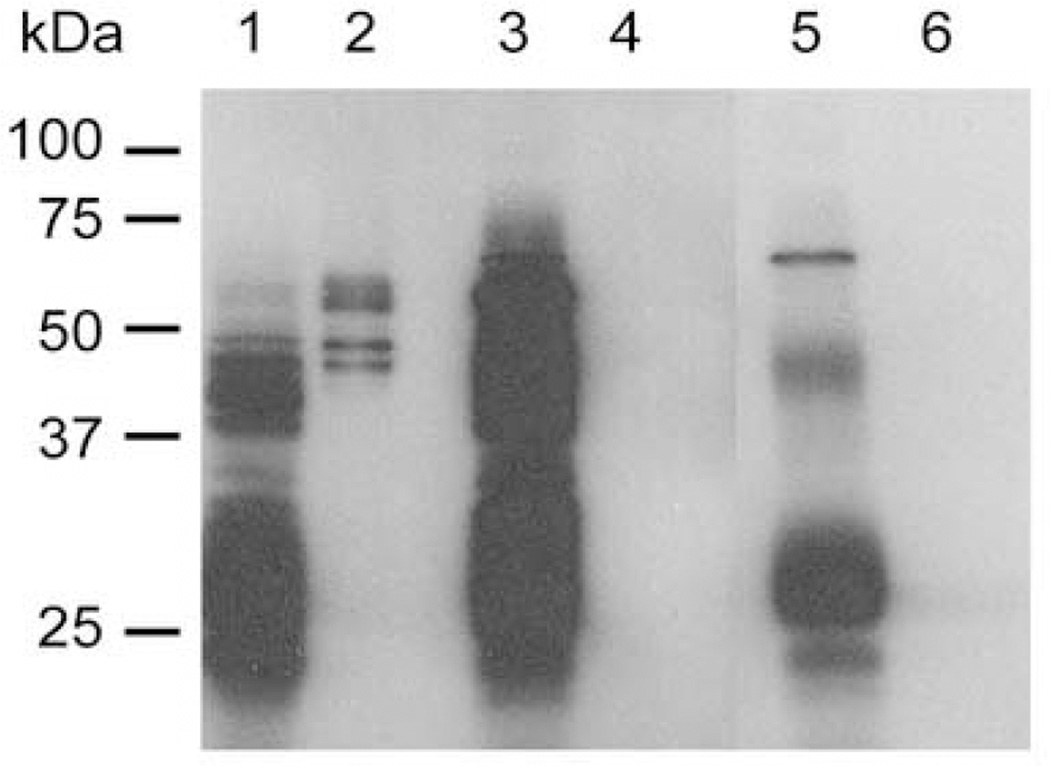
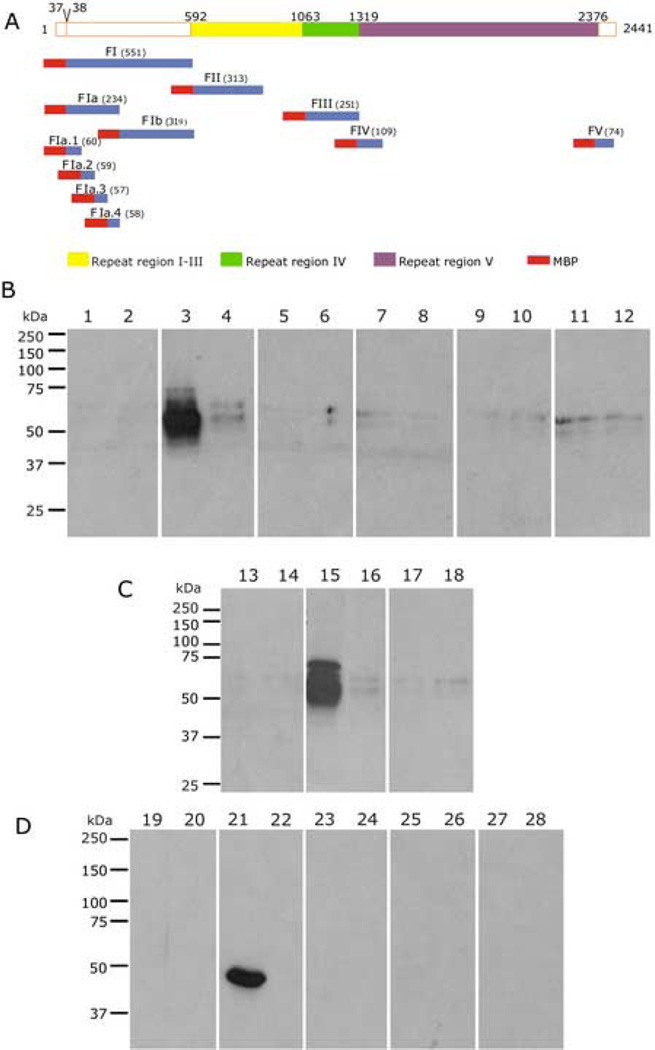
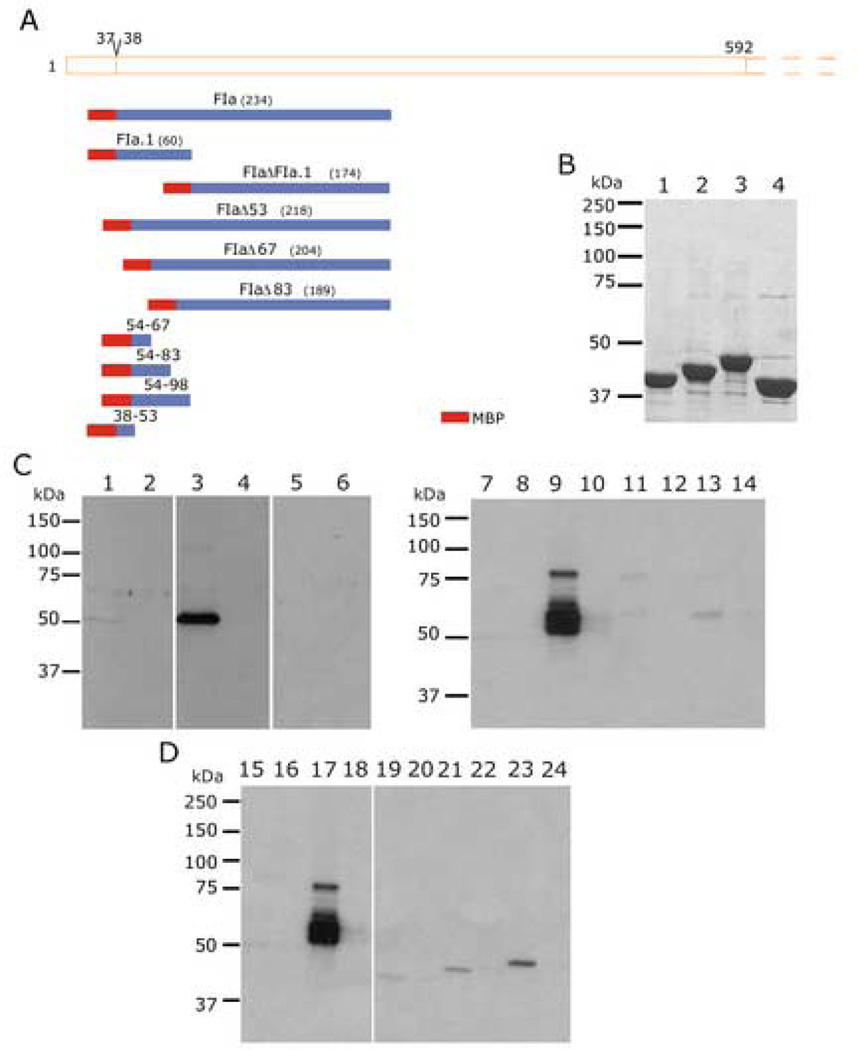
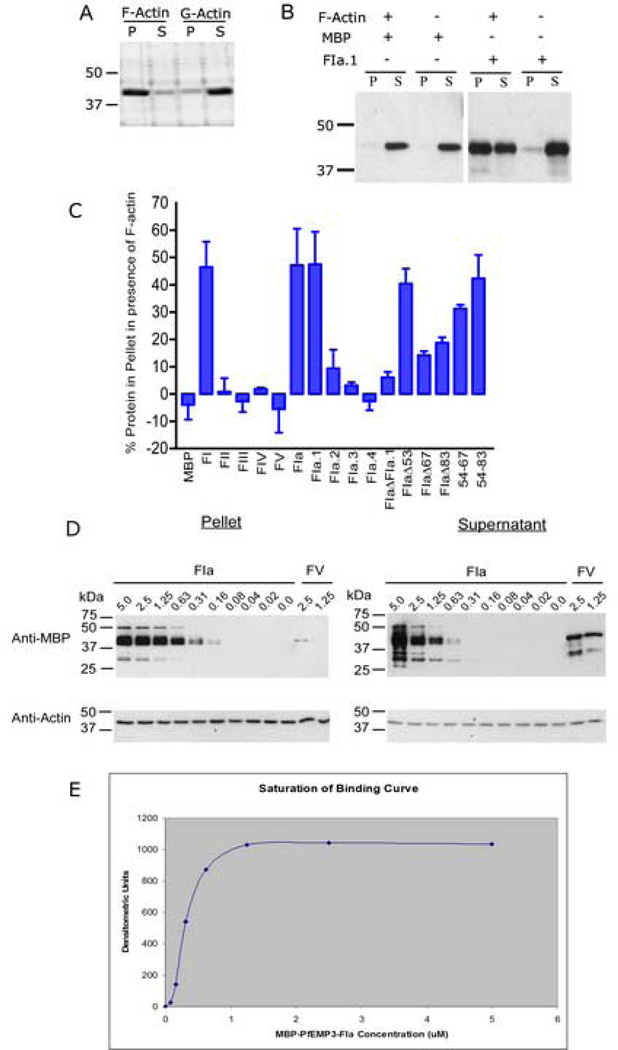
Plasmodium falciparum parasites express and traffick numerous proteins into the red blood cell (RBC), where some associate specifically with the membrane skeleton. Importantly, these interactions underlie the major alterations to the modified structural and functional properties of the parasite-infected RBC. P. falciparum Erythrocyte Membrane Protein 3 (PfEMP3) is one such parasite protein that is found in association with the membrane skeleton. Using recombinant PfEMP3 proteins in vitro, we have identified the region of PfEMP3 that binds to the RBC membrane skeleton, specifically to spectrin and actin. Kinetic studies revealed that residues 38-97 of PfEMP3 bound to purified spectrin with moderately high affinity (K(D(kin))=8.5 x 10(-8) M). Subsequent deletion mapping analysis further defined the binding domain to a 14-residue sequence (IFEIRLKRSLAQVL; K(D(kin))=3.8 x 10(-7) M). Interestingly, this same domain also bound to F-actin in a specific and saturable manner. These interactions are of physiological relevance as evidenced by the binding of this region to the membrane skeleton of inside-out RBCs and when introduced into resealed RBCs. Identification of a 14-residue region of PfEMP3 that binds to both spectrin and actin provides insight into the potential function of PfEMP3 in P. falciparum-infected RBCs.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
