

Restricted cell elongation in Arabidopsis hypocotyls is associated with a reduced average pectin esterification level
- PMID: 17572910
- PMCID: PMC1913053
- DOI: 10.1186/1471-2229-7-31
Restricted cell elongation in Arabidopsis hypocotyls is associated with a reduced average pectin esterification level
Abstract
Background: Cell elongation is mainly limited by the extensibility of the cell wall. Dicotyledonous primary (growing) cell walls contain cellulose, xyloglucan, pectin and proteins, but little is known about how each polymer class contributes to the cell wall mechanical properties that control extensibility.
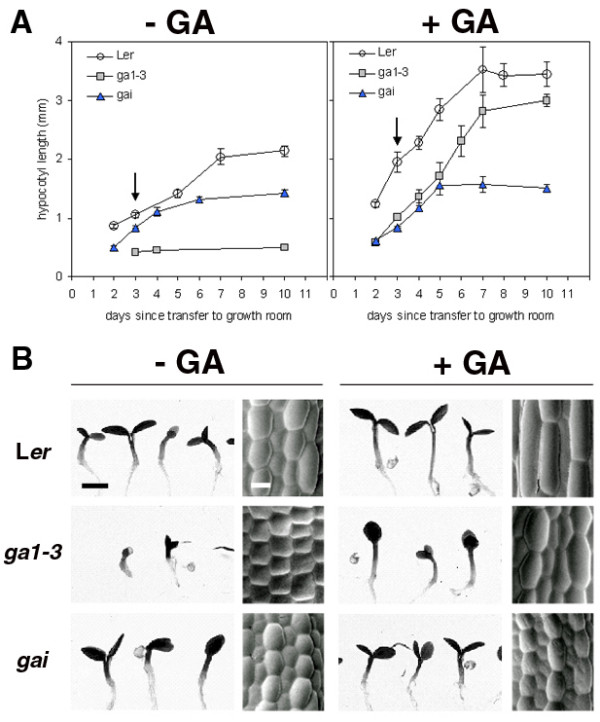
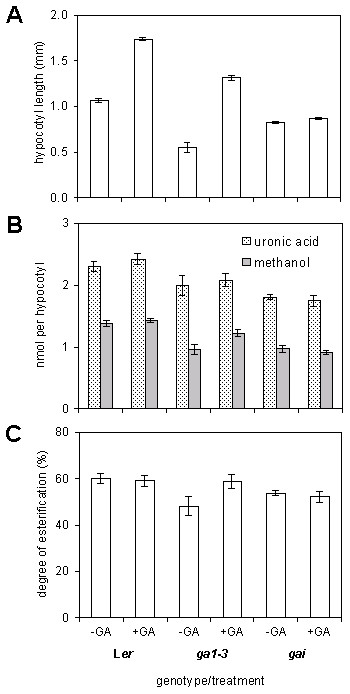
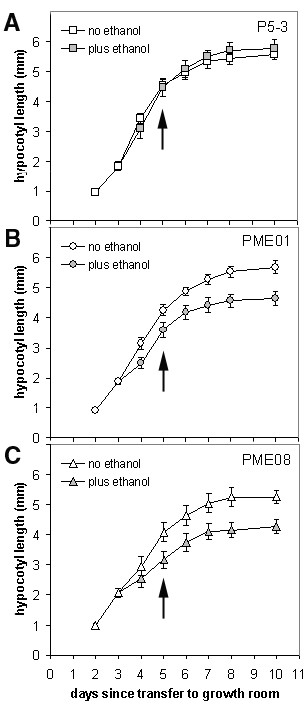
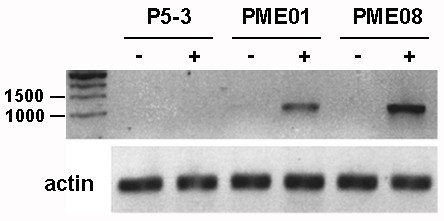
Results: We present evidence that the degree of pectin methyl-esterification (DE%) limits cell growth, and that a minimum level of about 60% DE is required for normal cell elongation in Arabidopsis hypocotyls. When the average DE% falls below this level, as in two gibberellic acid (GA) mutants ga1-3 and gai, and plants expressing pectin methyl-esterase (PME1) from Aspergillus aculeatus, then hypocotyl elongation is reduced.
Conclusion: Low average levels of pectin DE% are associated with reduced cell elongation, implicating PMEs, the enzymes that regulate DE%, in the cell elongation process and in responses to GA. At high average DE% other components of the cell wall limit GA-induced growth.
Figures
References
-
- McCann MC, Roberts K. Architecture of the primary cell wall. In: Lloyd C, editor. The Cytoskeletal Basis of Plant Growth and Form. London , Academic Press; 1991. pp. 109–129.
-
- Green PB. Organogenesis - a biophysical view. Annu Rev Plant Physiol Plant Molec Biol. 1980;31:51–82. doi: 10.1146/annurev.pp.31.060180.000411. - DOI
-
- Brummell DA, Lashbrook CC, Bennett AB. Plant endo-1,4-beta-D-glucanases - structure, properties, and physiological function. (Acs Symposium Series).Enzymatic Conversion of Biomass for Fuels Production. 1994;566:100–129.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
