

Rb regulates interactions between hematopoietic stem cells and their bone marrow microenvironment
- PMID: 17574022
- PMCID: PMC2768301
- DOI: 10.1016/j.cell.2007.03.055
Rb regulates interactions between hematopoietic stem cells and their bone marrow microenvironment
Abstract
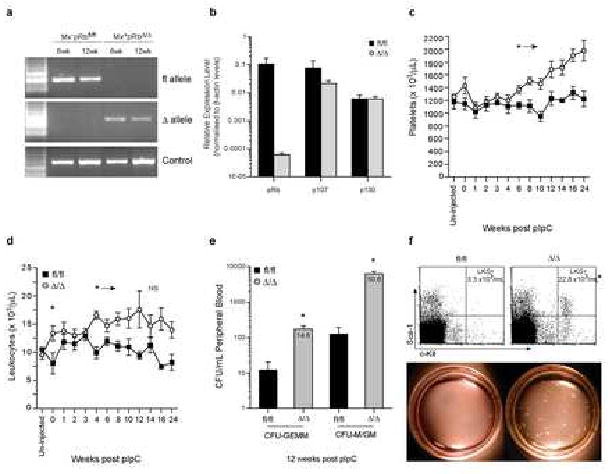
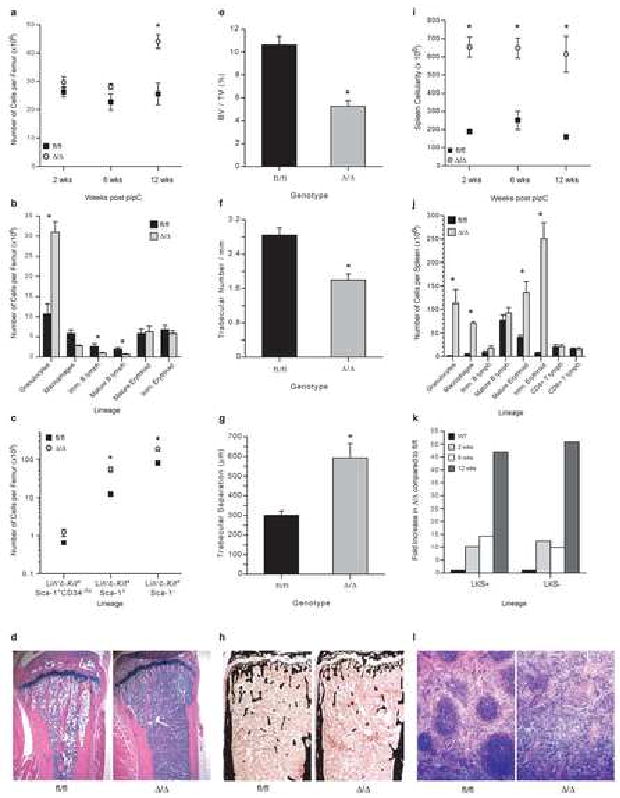
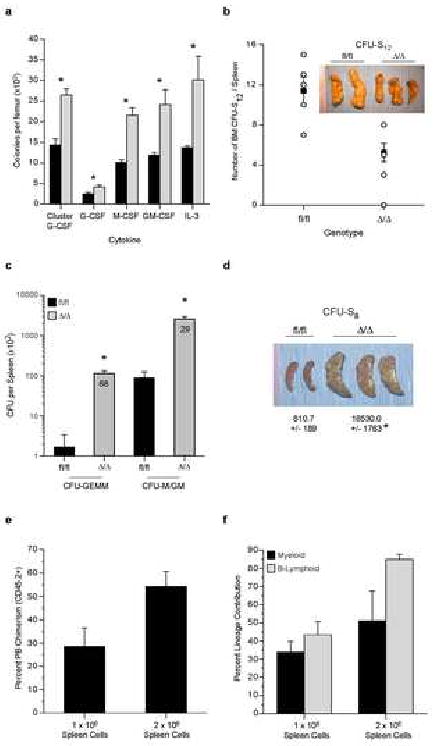
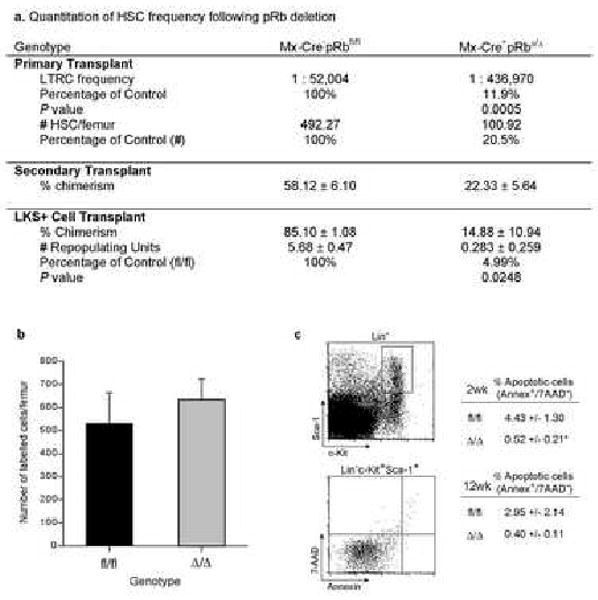
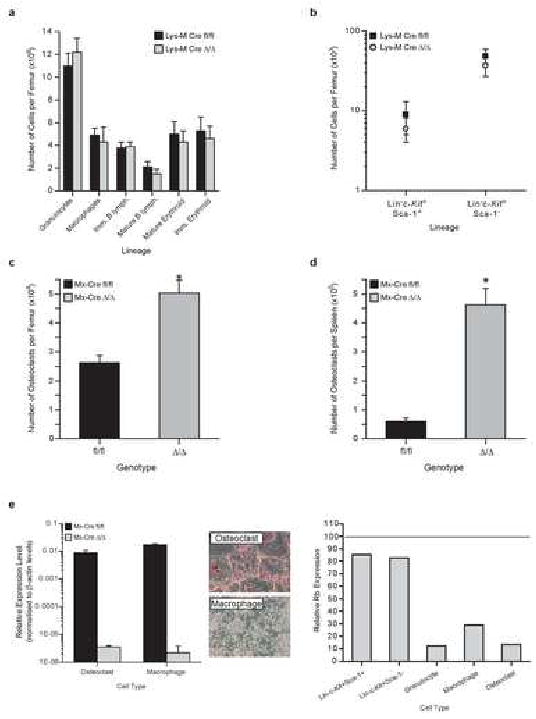
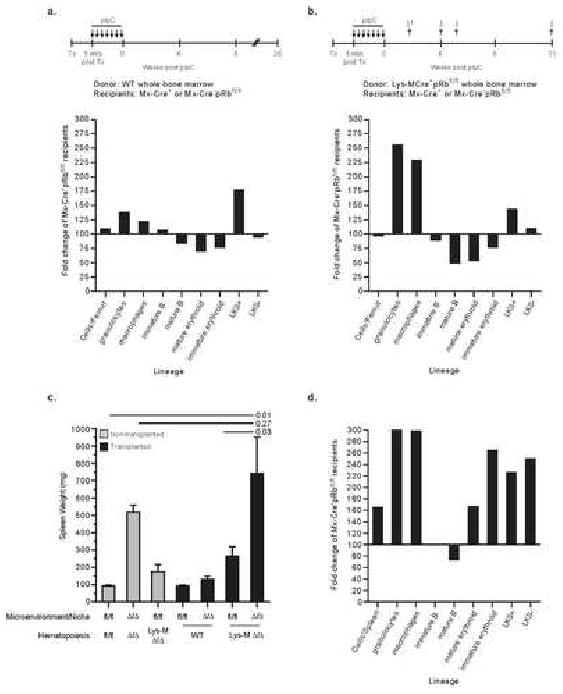
Hematopoiesis is maintained by stem cells (HSCs) that undergo fate decisions by integrating intrinsic and extrinsic signals, with the latter derived from the bone marrow (BM) microenvironment. Cell-cycle regulation can modulate stem cell fate, but it is unknown whether this represents an intrinsic or extrinsic effector of fate decisions. We have investigated the role of the retinoblastoma protein (RB), a central regulator of the cell cycle, in hematopoiesis. Widespread inactivation of RB in the murine hematopoietic system resulted in profound myeloproliferation. HSCs were lost from the BM due to mobilization to extramedullary sites and differentiation. This phenotype was not intrinsic to HSCs, but, rather, was the consequence of an RB-dependent interaction between myeloid-derived cells and the microenvironment. These findings demonstrate that myeloproliferation may result from perturbed interactions between hematopoietic cells and the niche. Therefore, RB extrinsically regulates HSCs by maintaining the capacity of the BM to support normal hematopoiesis and HSCs.
Conflict of interest statement
Conflict of Interest Statement: Authors have no conflicts of interest to declare pertaining to this research.
Figures
References
-
- Adams GB, Chabner KT, Alley IR, Olson DP, Szczepiorkowski ZM, Poznansky MC, Kos CH, Pollak MR, Brown EM, Scadden DT. Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor. Nature. 2006;439:599–603. - PubMed
-
- Adams GB, Scadden DT. The hematopoietic stem cell in its place. Nat Immunol. 2006;7:333–337. - PubMed
-
- Allinen M, Beroukhim R, Cai L, Brennan C, Lahti-Domenici J, Huang H, Porter D, Hu M, Chin L, Richardson A, et al. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell. 2004;6:17–32. - PubMed
-
- Anderson ARA, Weaver AM, Cummings PT, Quaranta V. Tumor Morphology and Phenotypic Evolution Driven by Selective Pressure from the Microenvironment. Cell. 2006;127:905–915. - PubMed
-
- Arai F, Hirao A, Ohmura M, Sato H, Matsuoka S, Takubo K, Ito K, Koh GY, Suda T. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118:149–161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
