

doi: 10.1101/gad.424807.
Positive feedback between Dia1, LARG, and RhoA regulates cell morphology and invasion
Affiliations
- PMID: 17575049
- PMCID: PMC1891425
- DOI: 10.1101/gad.424807
Item in Clipboard
Positive feedback between Dia1, LARG, and RhoA regulates cell morphology and invasion
Genes Dev.
.
Abstract
The RhoA-effector Dia1 controls actin-dependent processes such as cytokinesis, SRF transcriptional activity, and cell motility. Dia1 polymerizes actin through its formin homology (FH) 2 domain. Here we show that Dia1 acts upstream of RhoA independently of its effects on actin assembly. Dia1 binds to the leukemia-associated Rho-GEF (LARG) through RhoA-dependent release of Dia1 autoinhibition. The FH2 domain stimulates the guanine nucleotide exchange activity of LARG in vitro. Our results reveal that Dia1 is necessary for LPA-stimulated Rho/ROCK signaling and bleb-associated cancer cell invasion. Thus, Dia1-dependent RhoA activation constitutes a positive feedback mechanism to modulate cell behavior.
Figures
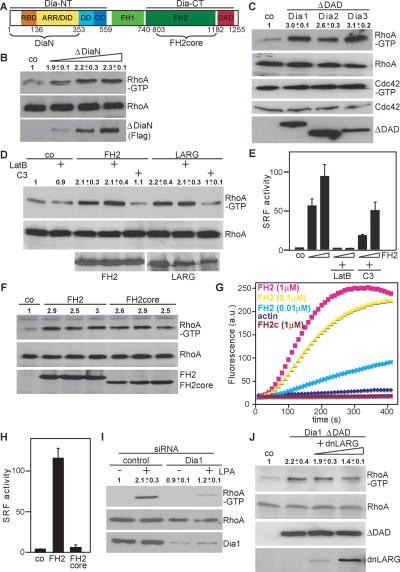
Role of Dia1 in activation of RhoA. (A) Schematic representation of mDia1. (B) Cells were transfected with control plasmid or increasing amounts of plasmids (0.2, 0.5, 1.0 μg) encoding ΔDiaN and subjected to RhoA-GTP pull-down assays. RhoA and Flag-ΔDiaN were analyzed by Western blotting. (C) Activation of RhoA but not Cdc42 by ΔDAD expression plasmids for Dia1–3 is shown. Active RhoA and Cdc42 were precipitated simultaneously from cell extracts. (D) FH2-induced RhoA activity in the absence or presence of 0.5 μM Latrunculin B (LatB) or in the absence or presence of C3 plasmid (0.5 μg). (E) Cells expressing FH2 (0.1 and 0.3 μg) together with the SRF reporter were cotransfected with C3 (0.1 μg) or pretreated with LatB as indicated. Reporter activity is shown as the mean ± SEM of three independent experiments. (F) FH2 or FH2core was transfected. The amount of Rho-GTP bound, of total RhoA, and of expressed FH2 and FH2core proteins was analyzed by Western blotting. (G) Shown is FH2-induced actin polymerizing versus FH2core capping activity using pyrene-actin assembly assays with indicated components. (H) Expression plasmids for FH2 or FH2core were transfected with an SRF reporter. Data represent the mean ± SEM of three independent experiments. (I) Dia1 is required for RhoA activation by LPA as shown by siRNA. Cells were untreated or stimulated with LPA (10 μM, 5 min) as indicated, and activated RhoA was precipitated. Cell extracts were immunoblotted for the indicated proteins. (J) HEK293 cells were transfected with Flag-Dia1ΔDAD and increasing amounts of expression plasmids encoding dnLARG (1.0, 5.0 μg). Activated RhoA and cell extracts were immunoblotted for the indicated proteins. All Western blot quantifications shown are the mean ± SEM of at least two independent experiments..
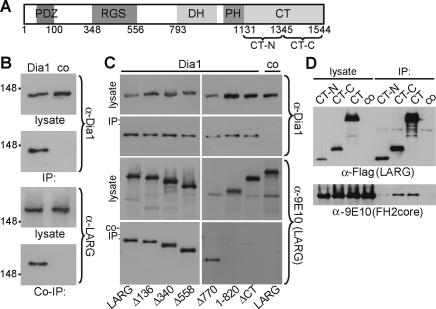
Dia1 interacts with LARG. (A) Schematic representation of LARG. (RGS) Regulator of G protein signaling; (DH) Dbl homology; (PH) pleckstrin homology. (B) Cells maintained in 10% serum were lysed before immunoprecipitation with α-Dia1 or control antiserum (co). Cell lysates and precipitates (IP) were analyzed by immunoblot as indicated. (C) Lysates were prepared from cells expressing myc-LARG derivatives or full-length LARG. Immunoprecipitates (IP) prepared using α-Dia1 were analyzed for associating LARG derivatives (Co-IP) by immunoblotting as indicated. (D) Lysates from cells expressing FH2core and LARG C-terminal mutants were immunoprecipitated and analyzed by immunoblotting as indicated.
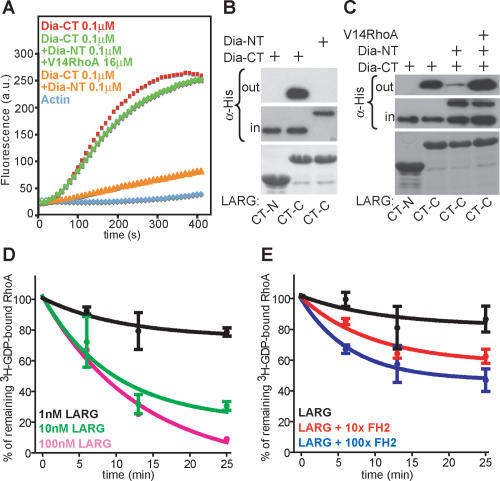
Dia1 stimulates LARG GEF activity. (A) Actin assembly assays of Dia1-CT show release of NT-induced inhibition by RhoAV14. (B) His-Dia-NT and His-Dia-CT proteins were incubated with GST-LARG C-terminal regions, and complexes were analyzed by immunoblot using α-His antibody. Coomassie staining for GST-LARG CT-N and CT-C is shown in the bottom panel. (C) GST pull-downs show association of LARG CT-C with Dia1-CT and the release of NT-induced binding inhibition by RhoAV14. Complexes were analyzed by immunoblot as indicated. Coomassie staining for GST-LARG CT-N and CT-C is shown in the bottom panel. One representative out of four independent experiments is shown. (D) Indicated concentrations of GST-LARG incubated with [8-3H]GDP-loaded RhoA. [8-3H]GDP-bound RhoA (%) is plotted against incubation time. Shown is the mean ± SEM of three independent experiments. (E) One nanomolar GST-LARG was incubated with [8-3H]GDP-loaded RhoA with 100-fold molar excess of GST or with 10- and 100-fold molar excess of GST-FH2. [8-3H]GDP-bound RhoA (%) is plotted against incubation time. Shown is the mean ± SEM of three independent experiments.
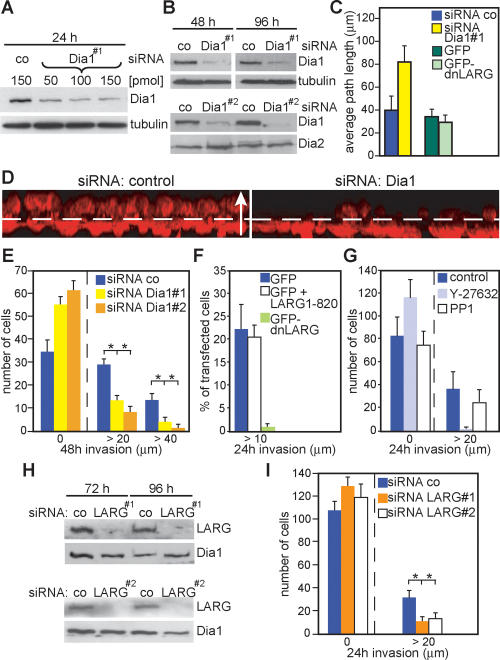
Dia1 is required for MDA-MB-435 cancer cell invasion. (A) Cells treated with different amounts of siRNA as indicated and analyzed by immunoblotting. (B) Cells treated with control siRNA or siRNA against Dia1 were immunoblotted for the indicated proteins. (C) Cells transfected with indicated siRNAs or plasmids were analyzed for random motility. Data are expressed as mean ± SD of three independent experiments. (D) Representative 3D reconstructions of invaded cells transfected with siRNA as indicated. Cells were stained for F-actin (red); dashed line indicates the approximate position of the Transwell membrane; the arrow (50 μm) indicates the direction of movement. (E) Cells were allowed to invade for 48 h into Matrigel before counting at indicated invasion distances. The average of cell number from three independent experiments ±SD is shown. (F) MDA-MB-435 cells transfected with indicated plasmids were analyzed in invasion assays. The percentage of GFP-positive cells invaded is shown from two independent experiments counting >300 cells. (G) The number of cells invaded into Matrigel containing LPA and indicated inhibitors is shown. (H,I) Cells treated with indicated siRNAs were analyzed for invasion.
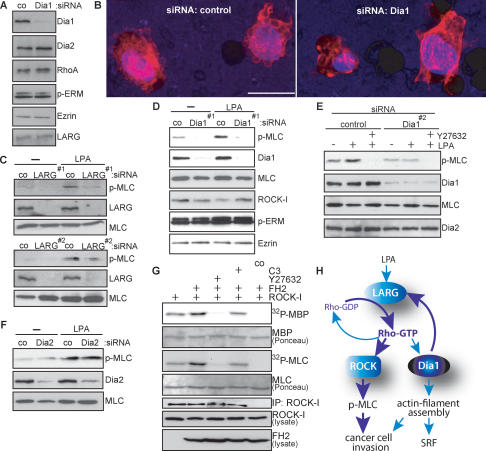
Dia1 is required for ROCK signaling and cancer cell morphology. (A) MDA-MB-435 cells were treated with control siRNA or siRNA against Dia1, and cell extracts were immunoblotted for the indicated proteins. (B) Representative 3D reconstructions of MDA-MB-435 cells treated in A invading through the porous membrane into the Matrigel. Cells were stained using DAPI (blue) to visualize nuclei and rhodamine-phalloidin (red) to visualize F-actin. Bar, 20 μm. (C–F) MDA-MB-435 cells treated with indicated siRNAs were stimulated with 10 μM LPA for 10 min. Cell extracts were immunoblotted for the indicated proteins. (G) myc-ROCK-I was immunoprecipitated from HEK293 cells cotransfected with Flag-FH2 or C3 or treated with 10 μM Y27632 as indicated, and subjected to kinase assays using MBP or MLC as substrates. Kinase reactions were separated by SDS-PAGE, and phosphorylated substrates were visualized by autoradiography (32P-MBP, 32P-MLC). Immobilized rabbit IgG was used as a control (co). One representative out of three independent experiments with similar results is shown. (H) Proposed hypothetical model for Dia1-mediated feedback activation of RhoA through LARG.
References
-
- Brandt D.T., Marion S., Griffiths G., Watanabe T., Kaibuchi K., Grosse R., Marion S., Griffiths G., Watanabe T., Kaibuchi K., Grosse R., Griffiths G., Watanabe T., Kaibuchi K., Grosse R., Watanabe T., Kaibuchi K., Grosse R., Kaibuchi K., Grosse R., Grosse R. Dia1 and IQGAP1 interact in cell migration and phagocytic cup formation. J. Cell Biol. 2007 (in press) - PMC - PubMed
-
- Chikumi H., Barac A., Behbahani B., Gao Y., Teramoto H., Zheng Y., Gutkind J.S., Barac A., Behbahani B., Gao Y., Teramoto H., Zheng Y., Gutkind J.S., Behbahani B., Gao Y., Teramoto H., Zheng Y., Gutkind J.S., Gao Y., Teramoto H., Zheng Y., Gutkind J.S., Teramoto H., Zheng Y., Gutkind J.S., Zheng Y., Gutkind J.S., Gutkind J.S. Homo- and hetero-oligomerization of PDZ-RhoGEF, LARG and p115RhoGEF by their C-terminal region regulates their in vivo Rho GEF activity and transforming potential. Oncogene. 2004;23:233–240. - PubMed
-
- Faix J., Grosse R., Grosse R. Staying in shape with formins. Dev. Cell. 2006;10:693–706. - PubMed
-
- Gomez T.S., Kumar K., Medeiros R.B., Shimizu Y., Leibson P.J., Billadeau D.D., Kumar K., Medeiros R.B., Shimizu Y., Leibson P.J., Billadeau D.D., Medeiros R.B., Shimizu Y., Leibson P.J., Billadeau D.D., Shimizu Y., Leibson P.J., Billadeau D.D., Leibson P.J., Billadeau D.D., Billadeau D.D. Formins regulate the actin-related protein 2/3 complex-independent polarization of the centrosome to the immunological synapse. Immunity. 2007;26:177–190. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous