

Nucleosome stability mediated by histone variants H3.3 and H2A.Z
- PMID: 17575053
- PMCID: PMC1891429
- DOI: 10.1101/gad.1547707
Nucleosome stability mediated by histone variants H3.3 and H2A.Z
Abstract
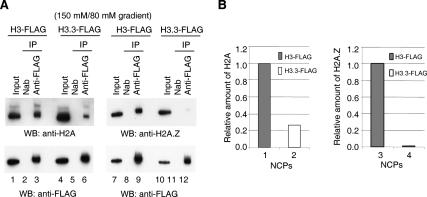
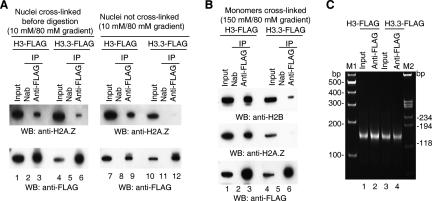
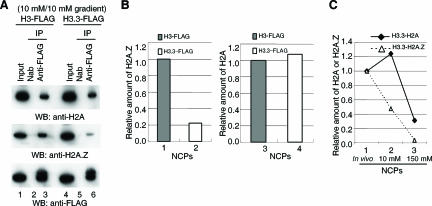
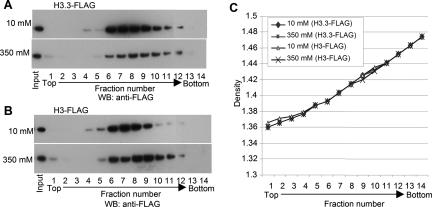
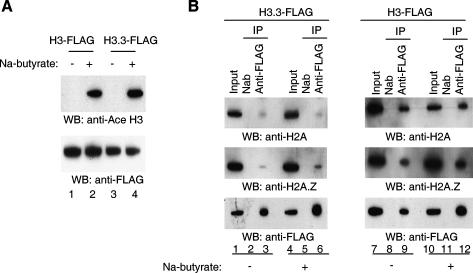
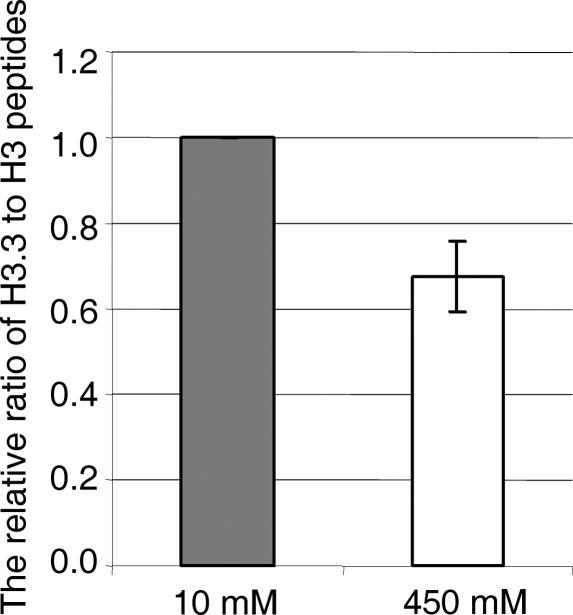
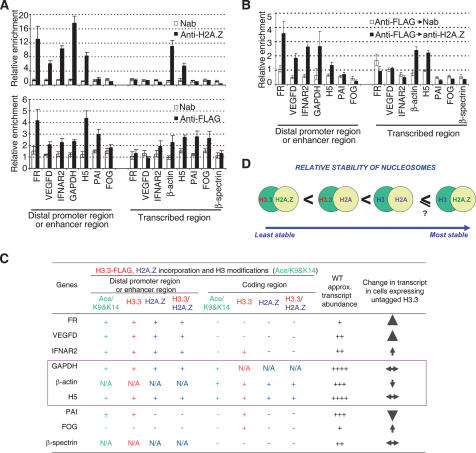
Nucleosomes containing the histone variant H3.3 tend to be clustered in vivo in the neighborhood of transcriptionally active genes and over regulatory elements. It has not been clear, however, whether H3.3-containing nucleosomes possess unique properties that would affect transcription. We report here that H3.3 nucleosomes isolated from vertebrates, regardless of whether they are partnered with H2A or H2A.Z, are unusually sensitive to salt-dependent disruption, losing H2A/H2B or H2A.Z/H2B dimers. Immunoprecipitation studies of nucleosome core particles (NCPs) show that NCPs that contain both H3.3 and H2A.Z are even less stable than NCPs containing H3.3 and H2A. Intriguingly, NCPs containing H3 and H2A.Z are at least as stable as H3/H2A NCPs. These results establish an hierarchy of stabilities for native nucleosomes carrying different complements of variants, and suggest how H2A.Z could play different roles depending on its partners within the NCP. They also are consistent with the idea that H3.3 plays an active role in maintaining accessible chromatin structures in enhancer regions and transcribed regions. Consistent with this idea, promoters and enhancers at transcriptionally active genes and coding regions at highly expressed genes have nucleosomes that simultaneously carry both H3.3 and H2A.Z, and should therefore be extremely sensitive to disruption.
Figures
References
-
- Abbott D.W., Ivanova V.S., Wang X., Bonner W.M., Ausio J., Ivanova V.S., Wang X., Bonner W.M., Ausio J., Wang X., Bonner W.M., Ausio J., Bonner W.M., Ausio J., Ausio J. Characterization of the stability and folding of H2A.Z chromatin particles: Implications for transcriptional activation. J. Biol. Chem. 2001;276:41945–41949. - PubMed
-
- Ahmad K., Henikoff S., Henikoff S. The histone variant H3.3 marks active chromatin by replication-independent nucleosome assembly. Mol. Cell. 2002;9:1191–1200. - PubMed
-
- Ashe H.L., Monks J., Wijgerde M., Fraser P., Proudfoot N.J., Monks J., Wijgerde M., Fraser P., Proudfoot N.J., Wijgerde M., Fraser P., Proudfoot N.J., Fraser P., Proudfoot N.J., Proudfoot N.J. Intergenic transcription and transinduction of the human β-globin locus. Genes & Dev. 1997;11:2494–2509. - PMC - PubMed
-
- Bruce K., Myers F.A., Mantouvalou E., Lefevre P., Greaves I., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Myers F.A., Mantouvalou E., Lefevre P., Greaves I., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Mantouvalou E., Lefevre P., Greaves I., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Lefevre P., Greaves I., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Greaves I., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Bonifer C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Tremethick D.J., Thorne A.W., Crane-Robinson C., Thorne A.W., Crane-Robinson C., Crane-Robinson C. The replacement histone H2A.Z in a hyperacetylated form is a feature of active genes in the chicken. Nucleic Acids Res. 2005;33:5633–5639. - PMC - PubMed
-
- Burton D.R., Butler M.J., Hyde J.E., Phillips D., Skidmore C.J., Walker I.O., Butler M.J., Hyde J.E., Phillips D., Skidmore C.J., Walker I.O., Hyde J.E., Phillips D., Skidmore C.J., Walker I.O., Phillips D., Skidmore C.J., Walker I.O., Skidmore C.J., Walker I.O., Walker I.O. The interaction of core histones with DNA: Equilibrium binding studies. Nucleic Acids Res. 1978;5:3643–3663. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources