

Nearest-neighbor non-additivity versus long-range non-additivity in TATA-box structure and its implications for TBP-binding mechanism
- PMID: 17576671
- PMCID: PMC1935006
- DOI: 10.1093/nar/gkm451
Nearest-neighbor non-additivity versus long-range non-additivity in TATA-box structure and its implications for TBP-binding mechanism
Abstract
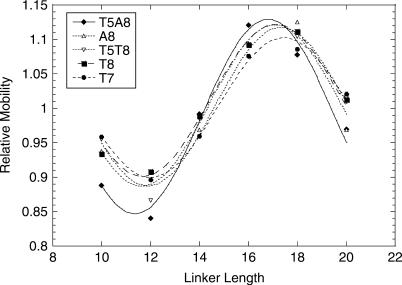
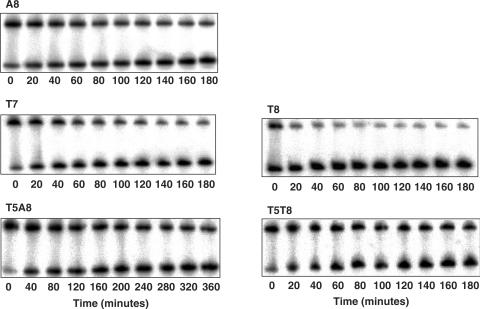
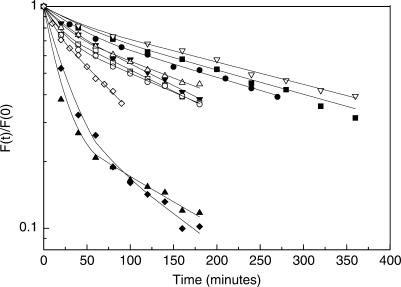
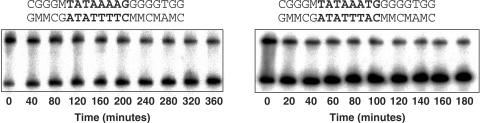
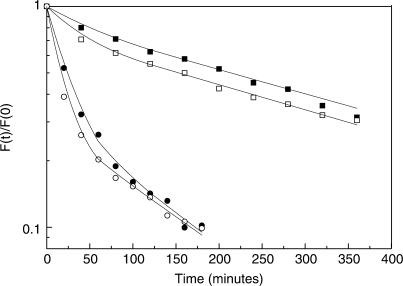
TBP recognizes its target sites, TATA boxes, by recognizing their sequence-dependent structure and flexibility. Studying this mode of TATA-box recognition, termed 'indirect readout', is important for elucidating the binding mechanism in this system, as well as for developing methods to locate new binding sites in genomic DNA. We determined the binding stability and TBP-induced TATA-box bending for consensus-like TATA boxes. In addition, we calculated the individual information score of all studied sequences. We show that various non-additive effects exist in TATA boxes, dependent on their structural properties. By several criterions, we divide TATA boxes to two main groups. The first group contains sequences with 3-4 consecutive adenines. Sequences in this group have a rigid context-independent cooperative structure, best described by a nearest-neighbor non-additive model. Sequences in the second group have a flexible, context-dependent conformation, which cannot be described by an additive model or by a nearest-neighbor non-additive model. Classifying TATA boxes by these and other structural rules clarifies the different recognition pathways and binding mechanisms used by TBP upon binding to different TATA boxes. We discuss the structural and evolutionary sources of the difficulties in predicting new binding sites by probabilistic weight-matrix methods for proteins in which indirect readout is dominant.
Figures
References
-
- Dickerson RE. Oxford Handbook of Nucleic Acid Structure. Oxford University Press; 1999. pp. 145–197.
-
- Travers AA. DNA-Protein: Structural interactions. Oxford: IRL Press; 1995. pp. 49–75.
-
- von Hippel PH. Biological Regulation and Development. Vol. 1. New York: Plenum Press; 1979. pp. 279–347.
-
- Kim JL, Nikolov DB, Burley SK. Co-crystal structure of TBP recognizing the minor groove of a TATA element. Nature. 1993;365:520–527. - PubMed
