

doi: 10.1083/jcb.200610108.
Essential and distinct roles for cdc42 and rac1 in the regulation of Schwann cell biology during peripheral nervous system development
Affiliations
- PMID: 17576798
- PMCID: PMC2064365
- DOI: 10.1083/jcb.200610108
Item in Clipboard
Essential and distinct roles for cdc42 and rac1 in the regulation of Schwann cell biology during peripheral nervous system development
J Cell Biol.
.
Abstract
During peripheral nervous system (PNS) myelination, Schwann cells must interpret extracellular cues to sense their environment and regulate their intrinsic developmental program accordingly. The pathways and mechanisms involved in this process are only partially understood. We use tissue-specific conditional gene targeting to show that members of the Rho GTPases, cdc42 and rac1, have different and essential roles in axon sorting by Schwann cells. Our results indicate that although cdc42 is required for normal Schwann cell proliferation, rac1 regulates Schwann cell process extension and stabilization, allowing efficient radial sorting of axon bundles.
Figures
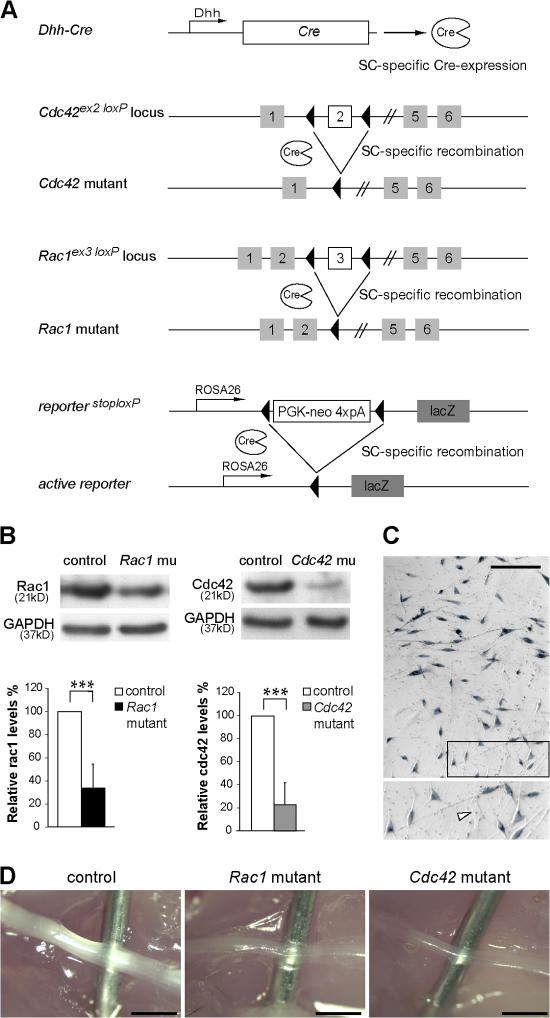
Recombination of the conditional Cdc42 and Rac1 alleles in SCs of mutant mice. (A) The regulatory sequences of the Dhh gene direct Cre recombinase (Cre) expression in SCs. Depicted are also the conditional Cdc42, Rac1, and the conditional reporter LacZ allele. Upon Dhh-Cre–mediated recombination, the genomic region between the two LoxP sites is excised, inactivating the Cdc42 and Rac1 genes. Recombination of the conditional LacZ reporter triggers the expression of β-galactosidase in SCs. (B) Western blot analysis shows a significant decrease in cdc42 and rac1 protein levels in P1 mutant sciatic nerve lysates (n = 5). ***, P < 0.001. Error bars indicate mean ± SD. (C) Widespread expression of β-galactosidase in SCs obtained from P0 nerves, as assessed by X-gal staining. The magnified box shows X-gal–positive SCs and an X-gal–negative fibroblast (arrowhead). (D) Sciatic nerves from P14 control, Cdc42 mutant, and Rac1 mutant mice. Note that both Cdc42 and Rac1 mutant sciatic nerves are thinner and more transparent than controls. For clarity, a needle is inserted behind the nerves. Bars, 1 mm.
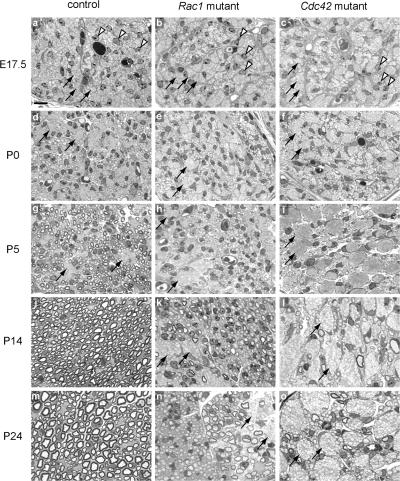
Axon sorting and myelination are impaired in Rac1 and Cdc42 mutant sciatic nerves. Semithin cross- sections of sciatic nerves from control (a, d, g, j, and m), Rac1 mutant (b, e, h, k, and n), and Cdc42 mutant (c, f, i, l, and o) mice stained with toluidine blue. In the controls, SCs (arrowheads) gradually segregate and myelinate single axons from the axon bundles (arrows). In contrast, in Rac1 and Cdc42 mutant nerves, axon sorting and myelination is impaired and axon bundles (arrows) persist. Although in P24 Rac1 mutant nerves, many axons are progressively sorted and myelinated, only very few axons are myelinated in P24 Cdc42 mutant nerves (o). Bar, 10 μm.
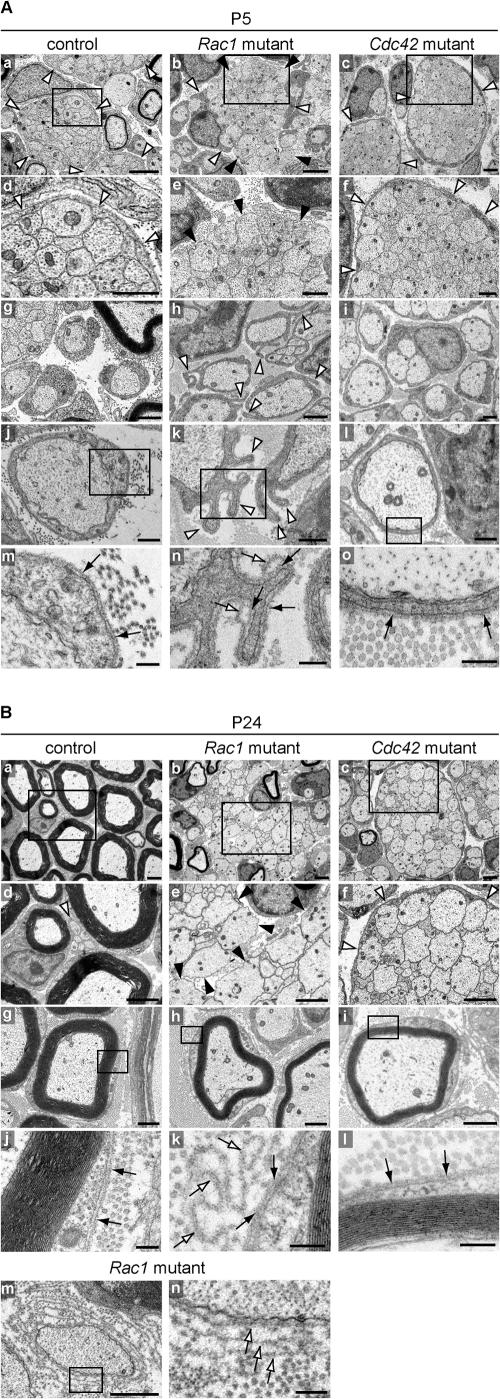
SC process extension and stabilization deficits in Rac1 mutant sciatic nerves. EM analysis of sciatic nerve cross-sections of control, Rac1 mutant, and Cdc42 mutant animals at P5 (A) and P24 (B). (A) At P5, immature SCs in control (a and d) and Cdc42 mutant (c and f) nerves extend long processes that fully envelop axon bundles (white arrowheads), a normal feature of SCs at this stage of differentiation. In contrast, in Rac1 mutant nerves, immature SC processes are shorter (b, white arrowheads) and often fail to envelope axon bundles (b and e, black arrowheads). Control (g and j) and Cdc42 mutant (i and l) promyelinating SCs are surrounded by an apposed BL (black arrows in m and o, respectively). In contrast, Rac1 mutant promyelinating SCs contain abnormal cytoplasmic protrusions that extend in various directions (h and k, white arrowheads). These protrusions are also surrounded by an apposed BL (n, black arrows); however, they often display empty BL loops above the apposed BL (n, white arrows). At the sites where they emerge, these BL loops are continuous with the apposed BL. Bars: (a–c) 2 μm; (d–i) 1 μm; (j–l) 500 nm; (m–o) 200 nm. (B) At P24, radial sorting in control nerves is virtually completed. While large caliber axons are being myelinated (a, d, and g), the small caliber axons are engaged by nonmyelinating SCs (d, white arrowhead). Myelinating SCs are surrounded by an apposed BL (j, black arrows). In P24 mutant nerves, hypomyelination is pronounced and resembles the situation at P5. Axon bundles in Rac1 mutant nerves are still not enveloped by immature SC processes (b and e, black arrowheads). Rac1 mutant myelinating SCs (h) do not show cytoplasmic protrusions. However, several loops of empty BL in the shape of the protrusions (k, white arrows) remain attached to the apposed BL (k, black arrows). In Rac1 mutant nerves, some naked axons are surrounded by several layers of BL, suggesting that processes of adjacent SCs repeatedly engaged and retracted from these axons, leaving behind empty BL layers (m and n, white arrows). P24 Cdc42 mutant nerves still contain many axon bundles (c), which as in P5 nerves (c and d, white arrowhead), are fully enveloped by immature SC processes (f, white arrowheads). P24 Cdc42 mutant nerves also contain myelinating SC-axon profiles (i), which are surrounded by an apposed BL (l, black arrows). Bars: (a–f) 2 μm; (g–i and m) 1 μm; (j–l and n) 200 nm.
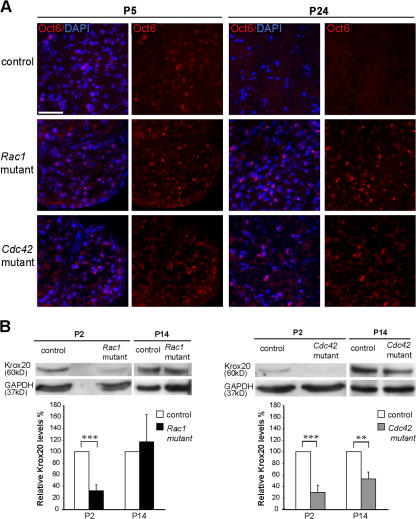
Altered Oct6 and Krox20 expression in Cdc42 and Rac1 mutant nerves. (A) Transverse sections of P5 and P24 sciatic nerves from control, Rac1 mutant, and Cdc42 mutant mice were stained for Oct6 (red) and DAPI (blue). Oct6-positive nuclei are present in P5 control, Rac1 mutant, and Cdc42 mutant nerves. At P24, Oct6 is still expressed in Rac1 and Cdc42 mutant nerves, whereas no Oct6-positive nuclei are present in control nerves. Bar, 100 μm. (B) Western blot analysis of sciatic nerve lysates of P2 Rac1 and Cdc42 mutant nerves show a significant decrease of Krox20 expression in relation to control. At P14, Krox20 levels were still significantly lower in Cdc42 mutant but not in Rac1 mutant nerves. Error bars indicate mean ± SD. **, P < 0.01; ***, P < 0.001.
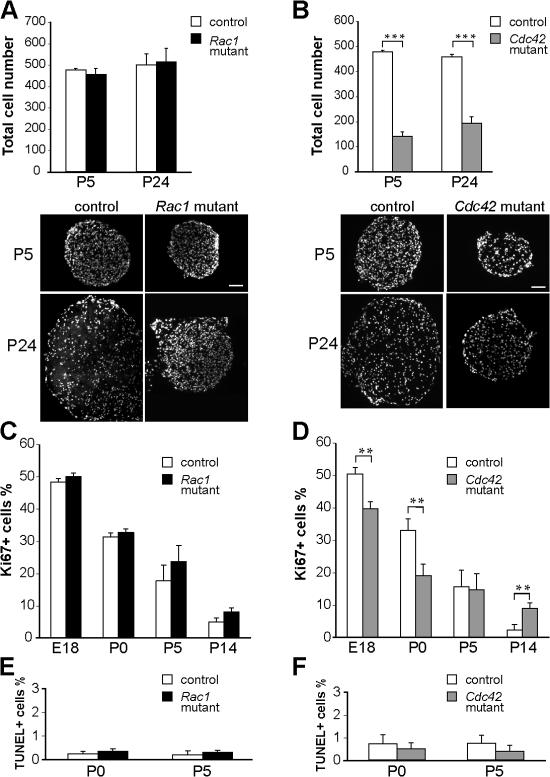
Total cell number, proliferation, and apoptosis in Rac1 and Cdc42 mutant sciatic nerves. (A) DAPI-stained P5 and P24 nerve cross-sections show similar numbers of cells in control and Rac1 mutant nerves but a significant decrease (B) of the total cell number in P5 and P24 Cdc42 mutant nerves. Transversal nerve sections obtained from mutant nerves are smaller than those of controls. ***, P < 0.001. (C) At E18, P0, P5, and P14, the numbers of proliferating Ki67-positive cells are not significantly different in sciatic nerves of control and Rac1 mutant mice. (D) In contrast, proliferation is significantly decreased in Cdc42 mutant nerves at E18 and P0 but significantly higher at P14. At P0 and P5, the number of apoptotic, TUNEL-positive cells is not significantly different in Rac1 mutant (E), Cdc42 mutant (F), and control nerves. Error bars indicate mean ± SD. **, P < 0.01. Bars, 100 μm.
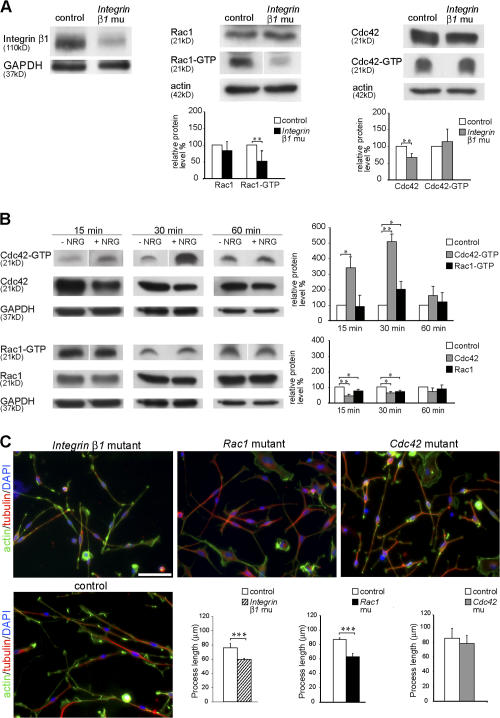
β1 integrin and NRG1 signaling regulate the activity of rac1 and cdc42. (A) Western blot analysis shows that β1 integrin protein expression is strongly reduced in P1 β1 integrin mutant (CNP-Cre
+ β1 integrinlox/0) sciatic nerve lysates compared with control (CNP-Cre
− β1 integrinlox/wt). Levels of active rac1 (rac1-GTP) and cdc42 (cdc42-GTP) were measured in a pull-down assay using GST-PAK-CD constructs. Nerve lysates from P5 β1 integrin mutant mice show significantly decreased levels of active rac1 compared with control lysates (n = 6). Activity of cdc42 is not significantly different between control and mutant lysates (n = 8). Total protein levels of cdc42 are significantly reduced. **, P < 0.01. (B) Exposure to 20 ng/ml NRG1 for 15 and 30 min significantly increases the levels of cdc42 activation in cultures of rat SCs compared with cultures of non–NRG1-treated SCs (control). Rac1 activity is only significantly higher than control after 30 min of exposure to NRG1. Note that the activity levels of cdc42 are increased severalfold in relation to those of rac1 after 15 and 30 min of NRG1 exposure. After 60 min of exposure to NRG1, both cdc42 and rac1 activities returned to control levels. After 15 or 30 min of NRG1 exposure, the total levels of cdc42 and rac1 are significantly reduced in relation to control. No significant differences were observed after 60 min. All experiments were repeated three times. *, P < 0.05; **, P < 0.01. (C) Acute SC cultures obtained from sciatic nerves of P1 β1 integrin, Rac1, and Cdc42 mutant and control mice were cultured for 16 h on PDL/laminin-2–coated dishes. The actin cytoskeleton (green), microtubules (red), and nuclei (blue) were visualized by phalloidin, tubulin antibodies, and DAPI, respectively. The mean length of SC processes is significantly decreased in β1 integrin and Rac1 mutant SC cultures but not in cdc42 mutant SCs (n = 5). Error bars indicate mean ± SD. ***, P < 0.001. Bar, 50 μm.
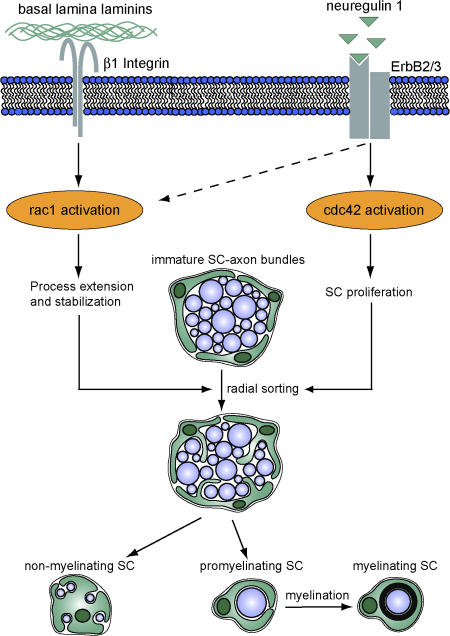
Proposed mechanism for the regulation of SC development by rac1 and cdc42. Cdc42 and rac1 play different but essential roles during SC development. Although rac1 can be activated by β1 integrin signaling and mediates SC process extension and stabilization, cdc42 is preferentially activated by NRG1 and is required for SC proliferation. Although rac1 and cdc42 play different roles in SC development, both are required for efficient radial sorting of axon bundles, a prerequisite for subsequent axon myelination.
Comment in
-
Myelination: all about Rac 'n' roll.J Cell Biol. 2007 Jun 18;177(6):953-5. doi: 10.1083/jcb.200705105. J Cell Biol. 2007. PMID: 17576794 Free PMC article. Review.
References
-
- Atanasoski, S., L. Notterpek, H.Y. Lee, F. Castagner, P. Young, M.U. Ehrengruber, D. Meijer, L. Sommer, E. Stavnezer, C. Colmenares, and U. Suter. 2004. The protooncogene Ski controls Schwann cell proliferation and myelination. Neuron. 43:499–511. - PubMed
-
- Benninger, Y., H. Colognato, T. Thurnherr, R.J. Franklin, D.P. Leone, S. Atanasoski, K.A. Nave, C. ffrench-Constant, U. Suter, and J.B. Relvas. 2006. Beta1-integrin signaling mediates premyelinating oligodendrocyte survival but is not required for CNS myelination and remyelination. J. Neurosci. 26:7665–7673. - PMC - PubMed
-
- Bradley, W.G., and M. Jenkison. 1975. Neural abnormalities in the dystrophic mouse. J. Neurol. Sci. 25:249–255. - PubMed
-
- Buttery, P.C., and C. ffrench-Constant. 1999. Laminin-2/integrin interactions enhance myelin membrane formation by oligodendrocytes. Mol. Cell. Neurosci. 14:199–212. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
