

TGF-beta signaling is essential for joint morphogenesis
- PMID: 17576802
- PMCID: PMC2064369
- DOI: 10.1083/jcb.200611031
TGF-beta signaling is essential for joint morphogenesis
Abstract
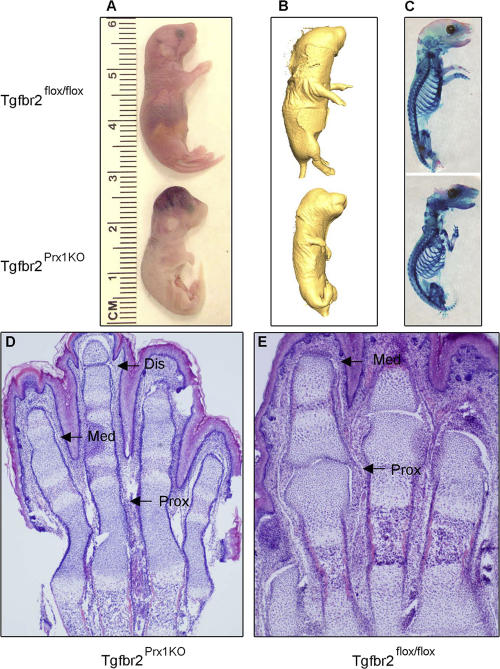
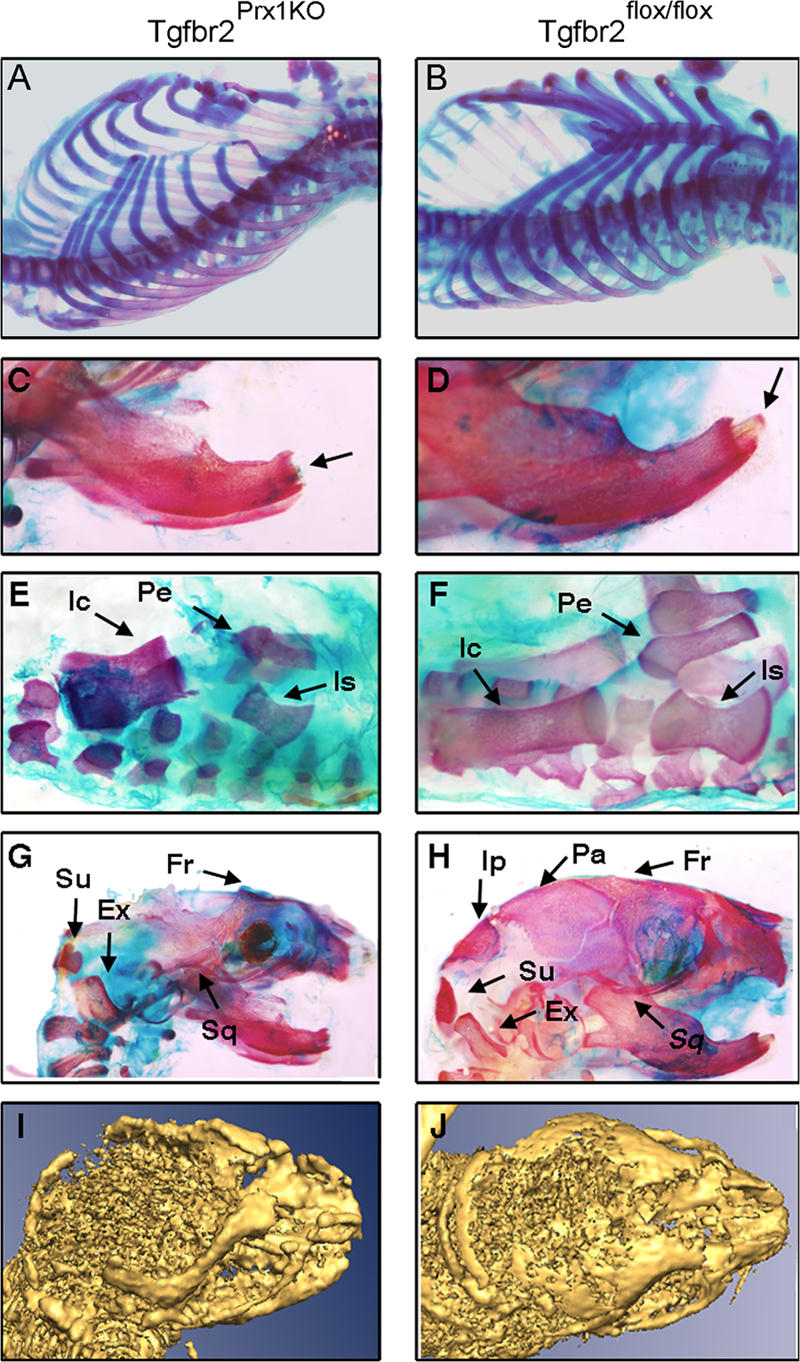
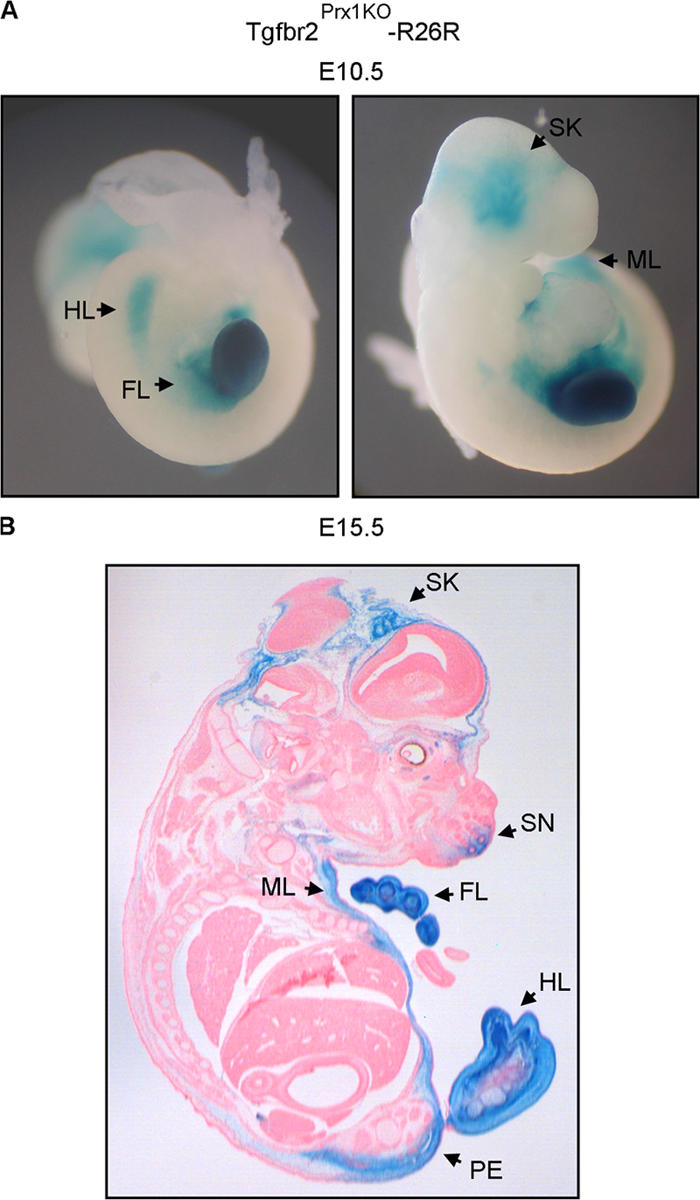
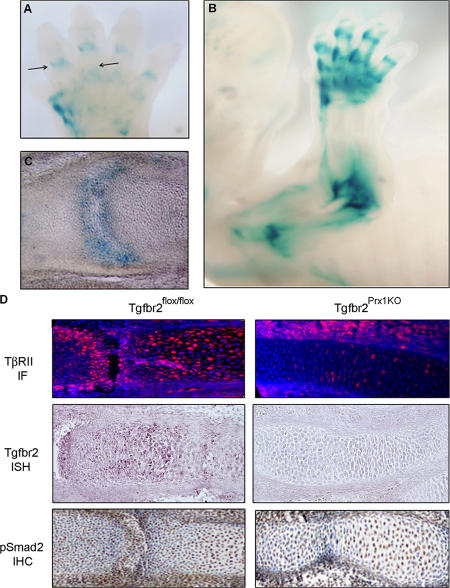
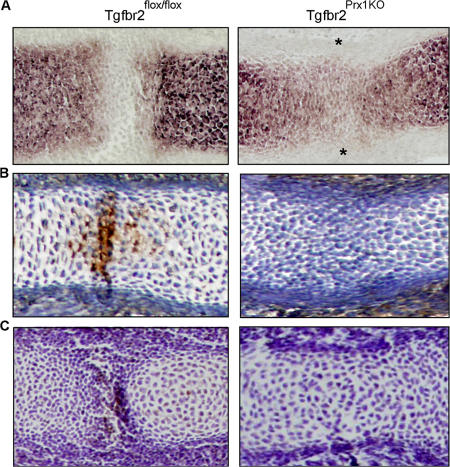
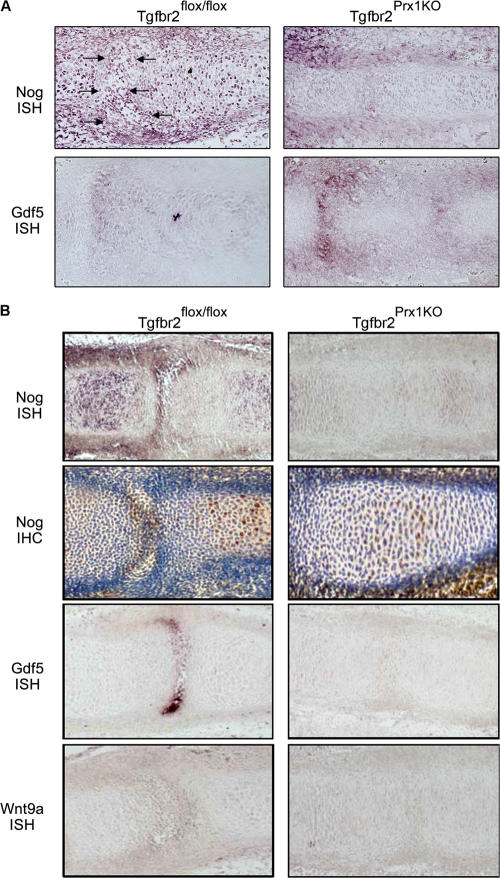
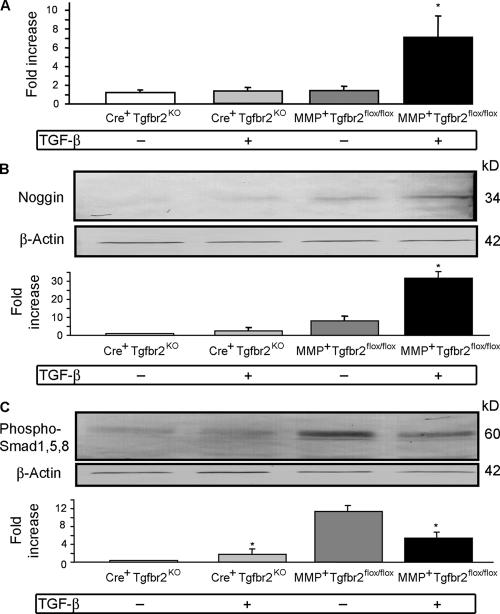
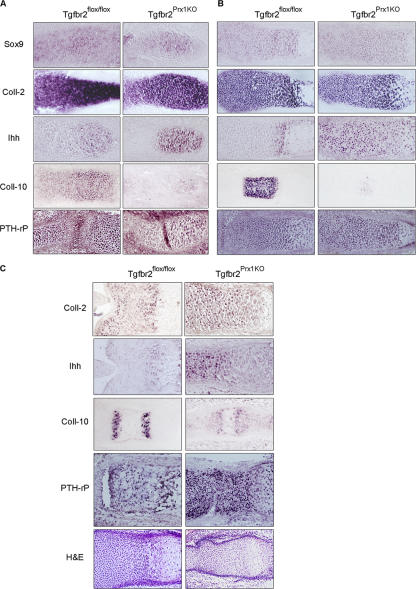
Despite its clinical significance, joint morphogenesis is still an obscure process. In this study, we determine the role of transforming growth factor beta (TGF-beta) signaling in mice lacking the TGF-beta type II receptor gene (Tgfbr2) in their limbs (Tgfbr2(PRX-1KO)). In Tgfbr2(PRX-1KO) mice, the loss of TGF-beta responsiveness resulted in the absence of interphalangeal joints. The Tgfbr2(Prx1KO) joint phenotype is similar to that in patients with symphalangism (SYM1-OMIM185800). By generating a Tgfbr2-green fluorescent protein-beta-GEO-bacterial artificial chromosome beta-galactosidase reporter transgenic mouse and by in situ hybridization and immunofluorescence, we determined that Tgfbr2 is highly and specifically expressed in developing joints. We demonstrated that in Tgfbr2(PRX-1KO) mice, the failure of joint interzone development resulted from an aberrant persistence of differentiated chondrocytes and failure of Jagged-1 expression. We found that TGF-beta receptor II signaling regulates Noggin, Wnt9a, and growth and differentiation factor-5 joint morphogenic gene expressions. In Tgfbr2(PRX-1KO) growth plates adjacent to interphalangeal joints, Indian hedgehog expression is increased, whereas Collagen 10 expression decreased. We propose a model for joint development in which TGF-beta signaling represents a means of entry to initiate the process.
Figures


References
-
- Ando, K., S. Kanazawa, T. Tetsuka, S. Ohta, X. Jiang, T. Tada, M. Kobayashi, N. Matsui, and T. Okamoto. 2003. Induction of Notch signaling by tumor necrosis factor in rheumatoid synovial fibroblasts. Oncogene. 22:7796–7803. - PubMed
-
- Archer, C.W., G.P. Dowthwaite, and P. Francis-West. 2003. Development of synovial joints. Birth Defects Res. C. Embryo Today. 69:144–155. - PubMed
-
- Baffi, M.O., E. Slattery, P. Sohn, H.L. Moses, A. Chytil, and R. Serra. 2004. Conditional deletion of the TGF-beta type II receptor in Col2a expressing cells results in defects in the axial skeleton without alterations in chondrocyte differentiation or embryonic development of long bones. Dev. Biol. 276:124–142. - PubMed
-
- Bhowmick, N.A., A. Chytil, D. Plieth, A.E. Gorska, N. Dumont, S. Shappell, M.K. Washington, E.G. Neilson, and H.L. Moses. 2004. TGF-beta signaling in fibroblasts modulates the oncogenic potential of adjacent epithelia. Science. 303:848–851. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
