

p75 neurotrophin receptor regulates tissue fibrosis through inhibition of plasminogen activation via a PDE4/cAMP/PKA pathway
- PMID: 17576803
- PMCID: PMC2064370
- DOI: 10.1083/jcb.200701040
p75 neurotrophin receptor regulates tissue fibrosis through inhibition of plasminogen activation via a PDE4/cAMP/PKA pathway
Abstract
Clearance of fibrin through proteolytic degradation is a critical step of matrix remodeling that contributes to tissue repair in a variety of pathological conditions, such as stroke, atherosclerosis, and pulmonary disease. However, the molecular mechanisms that regulate fibrin deposition are not known. Here, we report that the p75 neurotrophin receptor (p75(NTR)), a TNF receptor superfamily member up-regulated after tissue injury, blocks fibrinolysis by down-regulating the serine protease, tissue plasminogen activator (tPA), and up-regulating plasminogen activator inhibitor-1 (PAI-1). We have discovered a new mechanism in which phosphodiesterase PDE4A4/5 interacts with p75(NTR) to enhance cAMP degradation. The p75(NTR)-dependent down-regulation of cAMP results in a decrease in extracellular proteolytic activity. This mechanism is supported in vivo in p75(NTR)-deficient mice, which show increased proteolysis after sciatic nerve injury and lung fibrosis. Our results reveal a novel pathogenic mechanism by which p75(NTR) regulates degradation of cAMP and perpetuates scar formation after injury.
Figures


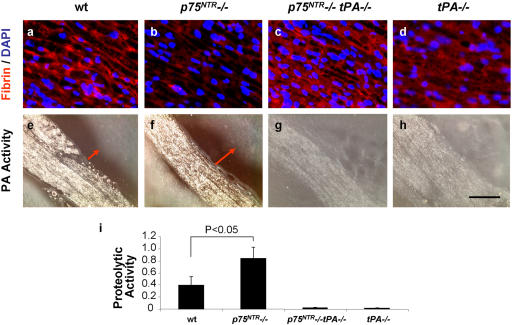







References
-
- Adams, R.A., M. Passino, B.D. Sachs, T. Nuriel, and K. Akassoglou. 2004. Fibrin mechanisms and functions in nervous system pathology. Mol. Interv. 4:163–176. - PubMed
-
- Akassoglou, K., W.-M. Yu, P. Akpinar, and S. Strickland. 2002. Fibrin inhibits peripheral nerve regeneration by arresting Schwann cell differentiation. Neuron. 33:861–875. - PubMed
-
- Allen, M.D., and J. Zhang. 2006. Subcellular dynamics of protein kinase A activity visualized by FRET-based reporters. Biochem. Biophys. Res. Commun. 348:716–721. - PubMed
-
- Barber, R., G.S. Baillie, R. Bergmann, M.C. Shepherd, R. Sepper, M.D. Houslay, and G.V. Heeke. 2004. Differential expression of PDE4 cAMP phosphodiesterase isoforms in inflammatory cells of smokers with COPD, smokers without COPD, and nonsmokers. Am. J. Physiol. Lung Cell. Mol. Physiol. 287:L332–L343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 5T32-GM07752/GM/NIGMS NIH HHS/United States
- P01 HD023315/HD/NICHD NIH HHS/United States
- NS21072/NS/NINDS NIH HHS/United States
- R01 NS021072/NS/NINDS NIH HHS/United States
- HD23315/HD/NICHD NIH HHS/United States
- NS27177/NS/NINDS NIH HHS/United States
- R01 NS052189/NS/NINDS NIH HHS/United States
- R01 NS027177/NS/NINDS NIH HHS/United States
- R01 NS051470/NS/NINDS NIH HHS/United States
- T32 GM007752/GM/NIGMS NIH HHS/United States
- R37 NS027177/NS/NINDS NIH HHS/United States
- NS51470/NS/NINDS NIH HHS/United States
- NS52189/NS/NINDS NIH HHS/United States
- R56 NS021072/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
