

BNIP3 is an RB/E2F target gene required for hypoxia-induced autophagy
- PMID: 17576813
- PMCID: PMC1952167
- DOI: 10.1128/MCB.02246-06
BNIP3 is an RB/E2F target gene required for hypoxia-induced autophagy
Abstract
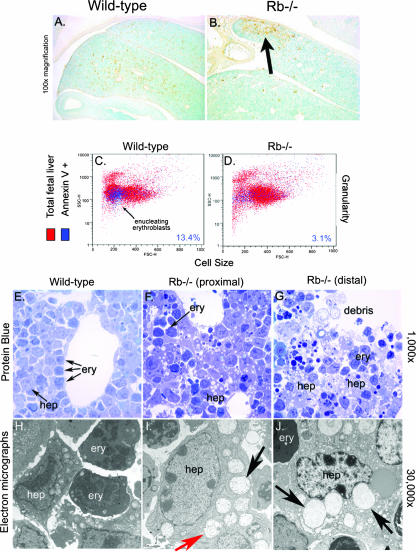
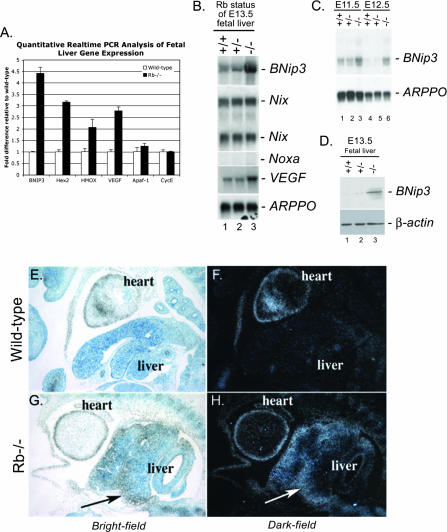
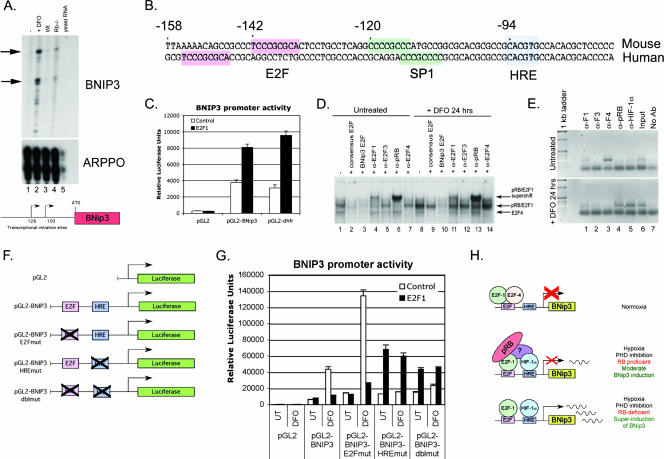
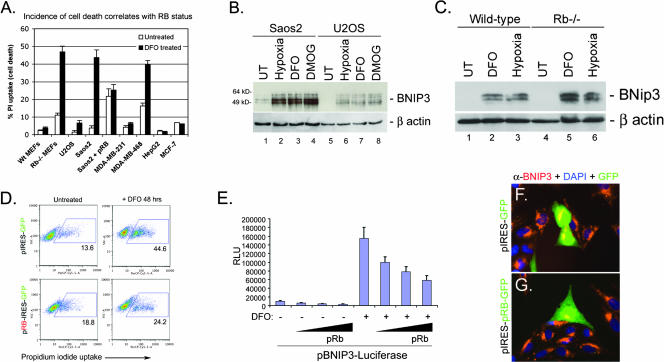
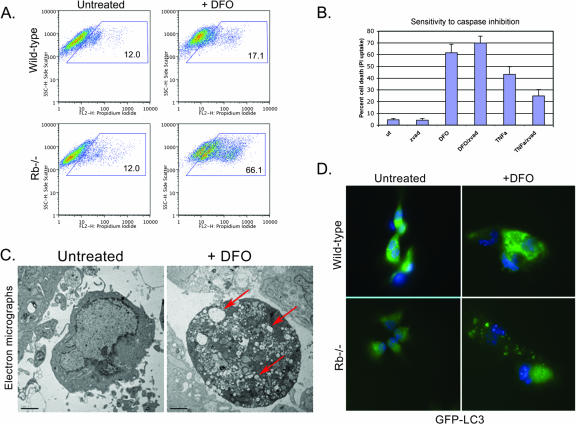
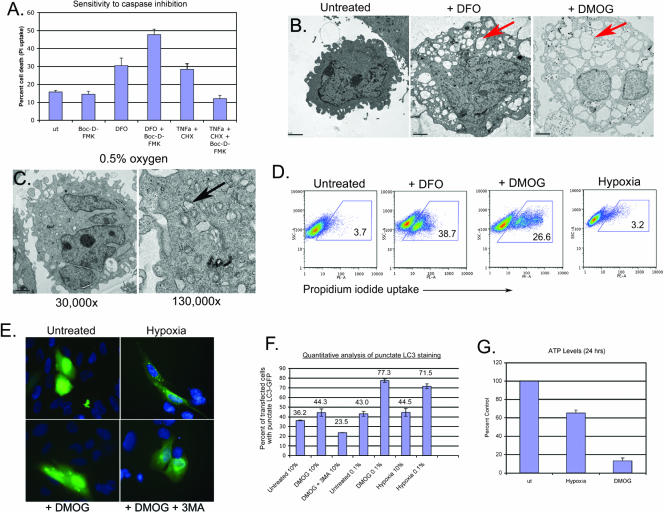
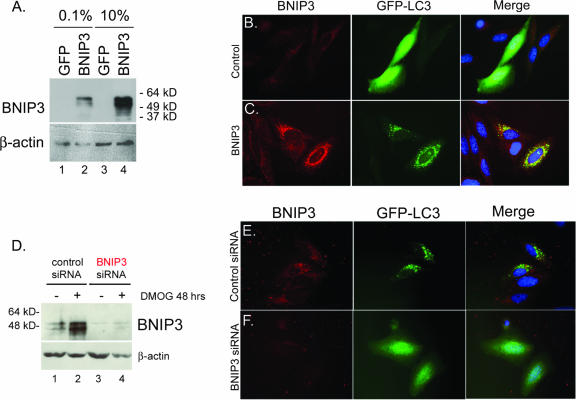
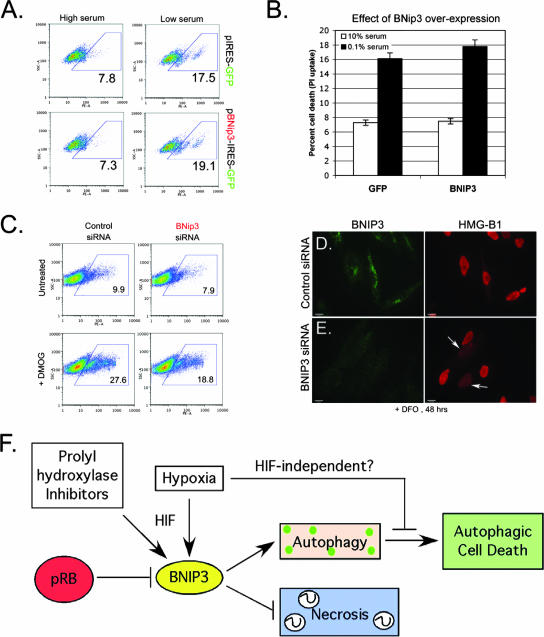
Hypoxia and nutrient deprivation are environmental stresses governing the survival and adaptation of tumor cells in vivo. We have identified a novel role for the Rb tumor suppressor in protecting against nonapoptotic cell death in the developing mouse fetal liver, in primary mouse embryonic fibroblasts, and in tumor cell lines. Loss of pRb resulted in derepression of BNip3, a hypoxia-inducible member of the Bcl-2 superfamily of cell death regulators. We identified BNIP3 as a direct target of pRB/E2F-mediated transcriptional repression and showed that pRB attenuates the induction of BNIP3 by hypoxia-inducible factor to prevent autophagic cell death. BNIP3 was essential for hypoxia-induced autophagy, and its ability to promote autophagosome formation was enhanced under conditions of nutrient deprivation. Knockdown of BNIP3 reduced cell death, and remaining deaths were necrotic in nature. These studies identify BNIP3 as a key regulator of hypoxia-induced autophagy and suggest a novel role for the RB tumor suppressor in preventing nonapoptotic cell death by limiting the extent of BNIP3 induction in cells.
Figures
References
-
- Bindra, R. S., S. L. Gibson, A. Meng, U. Westermark, M. Jasin, A. J. Pierce, R. G. Bristow, M. K. Classon, and P. M. Glazer. 2005. Hypoxia-induced down-regulation of BRCA1 expression by E2Fs. Cancer Res. 65:11597-11604. - PubMed
-
- Bindra, R. S., and P. M. Glazer. 2007. Repression of RAD51 gene expression by E2F4/p130 complexes in hypoxia. Oncogene 26:2048-2057. - PubMed
-
- Budde, A., N. Schneiderhan-Marra, G. Petersen, and B. Brune. 2005. Retinoblastoma susceptibility product pRB activates hypoxia-inducible factor (HIF-1). Oncogene 24:1802-1808. - PubMed
-
- Chau, B. N., and J. Y. J. Wang. 2003. Coordinated regulation of life and death by RB. Nat. Rev. Cancer 3:130-138. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases