

doi: 10.1101/gad.1561307.
Epub 2007 Jun 19.
Endosomal signaling of plant steroid receptor kinase BRI1
Affiliations
- PMID: 17578906
- PMCID: PMC1899468
- DOI: 10.1101/gad.1561307
Item in Clipboard
Endosomal signaling of plant steroid receptor kinase BRI1
Genes Dev.
.
Abstract
The LRR receptor serine/threonine kinases are a major eukaryotic receptor family, for which the central regulatory mechanism of endosomal trafficking remains largely unadressed. We show that the steroid receptor BRI1 localizes to both plasma membrane and early endosomal compartments, even when observed at low, endogenous expression levels, and that its localization and turnover are independent of ligand. However, increasing endosomal localization of BRI1 enhances activation of the pathway and genomic responses. Our data indicate distinct signaling and trafficking mechanisms within this receptor class and show that the use of endosomes as signaling compartments is an unexpectedly broad phenomenon in eukaryotes.
Figures
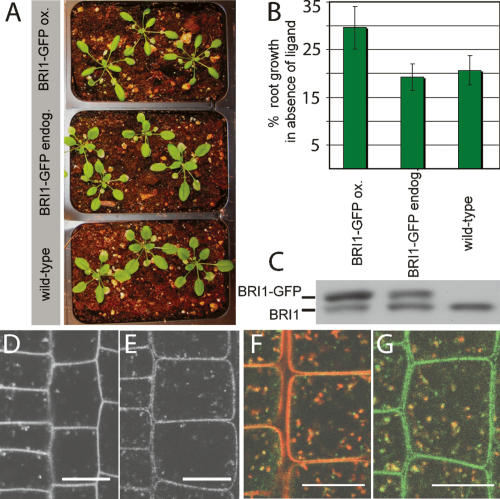
Endogenously expressed BRI1-GFP localizes to endosomes. (A) Representative pictures of rosette stage Arabidopsis grown under identical conditions. The BRI1-GFP line expressing at endogenous levels (endog.) is indistinguishable from wild type, whereas the overexpressing line (ox.) shows the reported overexpression phenotypes of narrow leaf blades and elongated, twisting petioles (leaf stalks). (B) Roots were depleted of endogenous BRs by growth on 5 μM brassinazole for 3 d. Primary root growth was assessed after three more days (n = 10 per line). Percent growth relative to untreated control is shown. The line expressing BRI1-GFP at endogenous levels is inhibited to wild-type levels, whereas the overexpressing line (ox.) shows less inhibition of root growth in the absence of ligand. (C) Immunoblot of the same lines detected with α-BRI1 antibodies. Intensity of the BRI1-GFP (top) band is much stronger than the band of endogenous BRI1 (bottom) in the overexpressing line, but both bands show the same intensity in middle lane. (D,E) Subcellular localization and levels of BRI1-GFP in root meristem epidermal cells. (D) BRI1-GFP-overexpressing line. (E) BRI1-GFP endogenous expresser. (F) BRI1-GFP (green) partially colocalizes with the endocytic tracer FM4-64 (red) after 5–10 min uptake. (G) BRI1-GFP (green) also colocalizes partially with VHA-a1-RFP (red). See Supplementary Figure 1 for individual channels of overlays. Bars, 10 μm.

Ligand-independent localization and turnover of BRI1. (A) Seedling roots were treated with saturating amounts of ligand (BL, 1 μM), after recording of first frame (0 min) and were observed at 1-min intervals. BRI1-GFP localization remains unaltered. (B) As in A, but roots were depleted of endogenous BR by growing on 5-μM brassinazole for 3 d. BRI1-GFP localization remains unchanged. (C) In vivo pulse-chase analysis of BRI1-YFP. Transgenic lines driving BRI1-YFP under a heat-shock-inducible promoter were used to measure the half-life of BRI1 in vivo and exclusively in root meristematic cells by quantitative confocal microscopy. YFP signal intensity peaked 4 h after heat shock, when mRNA levels were already back to basal levels (not shown). Turnover was recorded as the reduction in YFP intensity relative to peak expression. Data points were recorded every 2 min. Half-life of BRI1 was determined to be ∼5 h and this turnover rate appeared to be entirely independent of ligand, as much in untreated roots (left) as in ligand-depleted roots (right) (grown as in B). (Below) Picture row shows an example of signal recordings for five untreated control root meristems. (D, left) BKI-YFP displays clear PM accumulation over cytosolic background, whereas no comparable signal can be observed in endosomal compartments. (Right) BRI1-GFP grown under identical conditions. Bars: A,B, 10 μm.
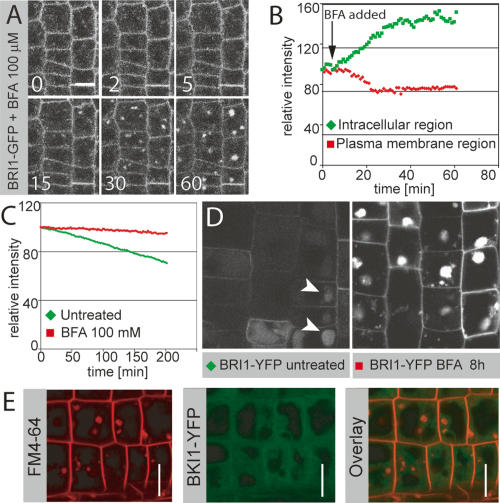
BFA increases endosomal localization of BRI1. (A) Time-lapse analysis of BFA-induced shift of BRI1 from the plasma membrane to endosomal compartments. (B) Quantification of time lapse in A. BFA induces a rapid drop in plasma membrane signal with a concomitant increase in intracellular fluorescence. A new steady state is apparently reached after ∼30 min. (C) Quantification of the effect of BFA on BRI1-YFP degradation in a pulse-chase experiment. BFA leads to a nearly complete block of BRI1 turnover. (D) BRI1-YFP localization in seedling root meristems 8 h after heat-shock-induced peak expression, with (left) or without (right) BFA treatment, taken under identical settings. BFA leads to strong accumulation of BRI1-YFP in endosomal aggregates, whereas only a weak and predominantly vacuolar BRI1-YFP signal is observed in the untreated control (arrowheads). (E) FM4-64 (red, left) and BKI1-YFP (green, middle) after 50 μM BFA treatment for 30 min. (Right) Overlay. Note the absence of BKI1 signal in BFA compartments. Bars: A,D,E, 10 μm.
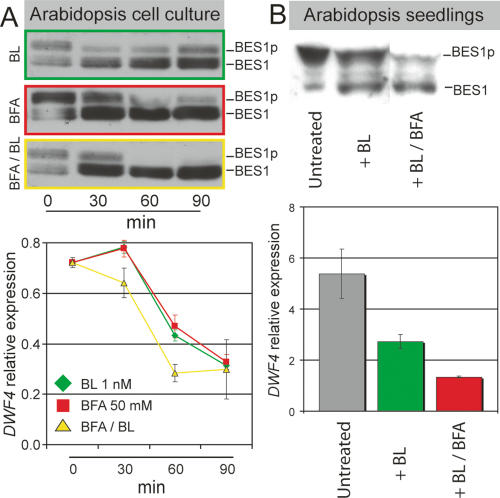
BRI1 signaling is induced by BFA treatment. (A) A highly sensitive Arabidopsis green cell suspension line shows dephosphorylation of BES1 (green, above) and suppression of early-response gene, DWF4 (green, below) already at 1 nM BL for 30 min. (Middle) BFA treatment alone leads to an efficient activation of BES1 (red) and suppression early-response genes. External BL enhances the BFA effect on BES1 (yellow, above) and DWF4 (yellow, below). (B) Enhancement of BR signaling by BFA can also be observed in BRI1-overexpressing seedlings, when treating with low amounts of BL (10 nM) for 4 h, which leads to only half-maximal dephosporylation of BES1 (above) and a twofold reduction in DWF4 levels (green, below). Adding BFA (100 μM) after 30 min of BL pretreatment significantly enhances BES1 activation (above) and DWF4 suppression (red, below).
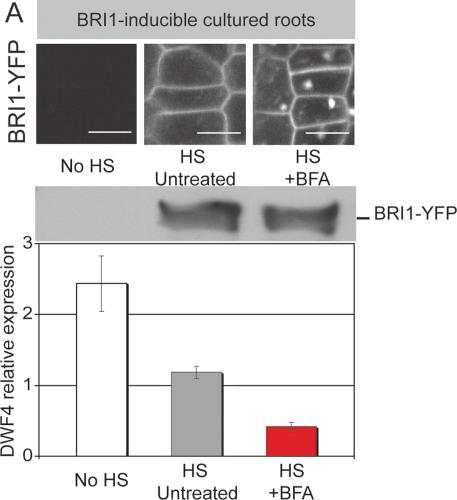
Induced BRI-YFP expression in cultured roots. (A) BFA effects can also be observed in intact organs using a bri1-null mutant root culture containing heat-shock-inducible BRI1-YFP. In this system, the effects of BFA on localization and signaling can be simultaneously assessed. Top panel shows BRI-YFP signals, 8 h after a 30-min heat-shock or control treatment. BFA treatment (100 μM) started 4 h after heat shock. Middle panel shows BRI1-YFP levels as detected on immunoblot with α-BRI1 after same treatments as above. Bottom panel shows heat-shock-induced suppression of DWF4 (gray) and its further reduction by BFA treatment (red). Bar, 10 μm.
Comment in
-
Signaling from plant endosomes: compartments with something to say!Genes Dev. 2007 Jul 1;21(13):1578-80. doi: 10.1101/gad.1577607. Genes Dev. 2007. PMID: 17606636 No abstract available.
References
-
- Ando R., Mizuno H., Miyawaki A., Mizuno H., Miyawaki A., Miyawaki A. Regulated fast nucleocytoplasmic shuttling observed by reversible protein highlighting. Science. 2004;306:1370–1373. - PubMed
-
- Baass P.C., Di Guglielmo G.M., Authier F., Posner B.I., Bergeron J.J., Di Guglielmo G.M., Authier F., Posner B.I., Bergeron J.J., Authier F., Posner B.I., Bergeron J.J., Posner B.I., Bergeron J.J., Bergeron J.J. Compartmentalized signal transduction by receptor tyrosine kinases. Trends Cell Biol. 1995;5:465–470. - PubMed
-
- Belkhadir Y., Chory J., Chory J. Brassinosteroid signaling: A paradigm for steroid hormone signaling from the cell surface. Science. 2006;314:1410–1411. - PubMed
-
- DeFea K.A., Zalevsky J., Thoma M.S., Dery O., Mullins R.D., Bunnett N.W., Zalevsky J., Thoma M.S., Dery O., Mullins R.D., Bunnett N.W., Thoma M.S., Dery O., Mullins R.D., Bunnett N.W., Dery O., Mullins R.D., Bunnett N.W., Mullins R.D., Bunnett N.W., Bunnett N.W. β-Arrestin-dependent endocytosis of proteinase-activated receptor 2 is required for intracellular targeting of activated ERK1/2. J. Cell Biol. 2000;148:1267–1281. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases