

C-reactive protein collaborates with plasma lectins to boost immune response against bacteria
- PMID: 17581635
- PMCID: PMC1933394
- DOI: 10.1038/sj.emboj.7601762
C-reactive protein collaborates with plasma lectins to boost immune response against bacteria
Abstract
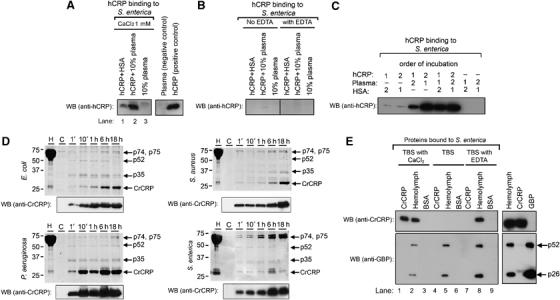
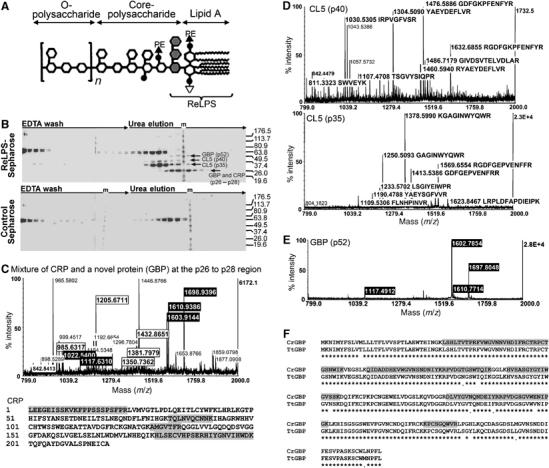
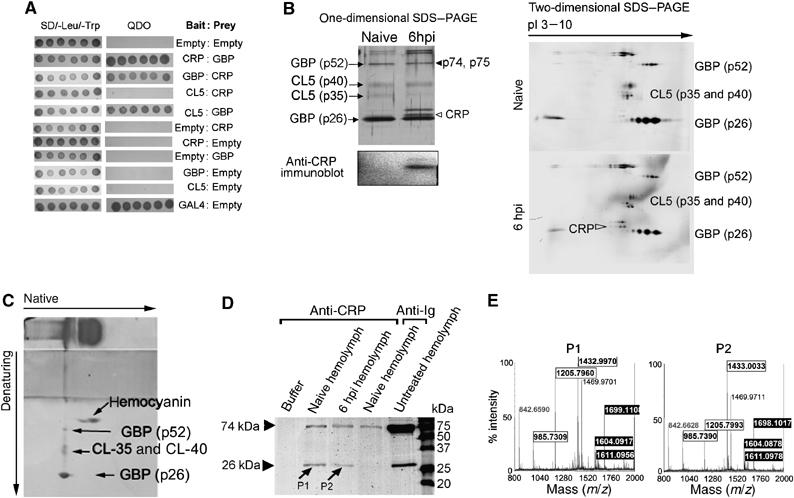
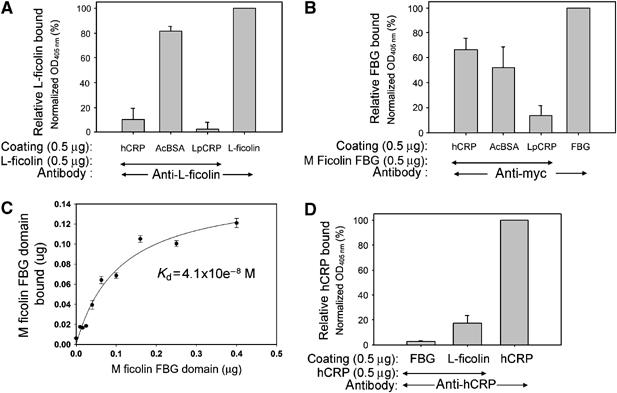
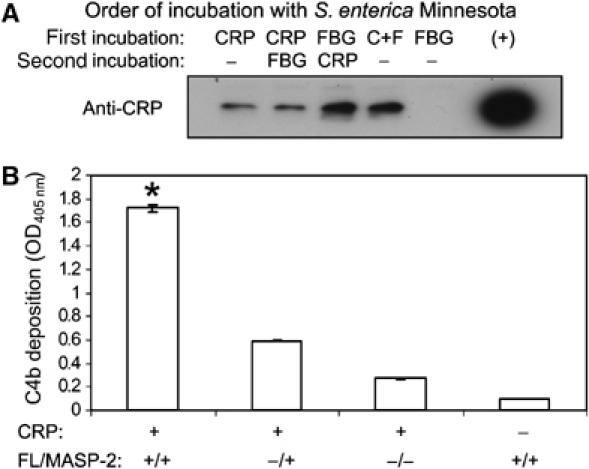
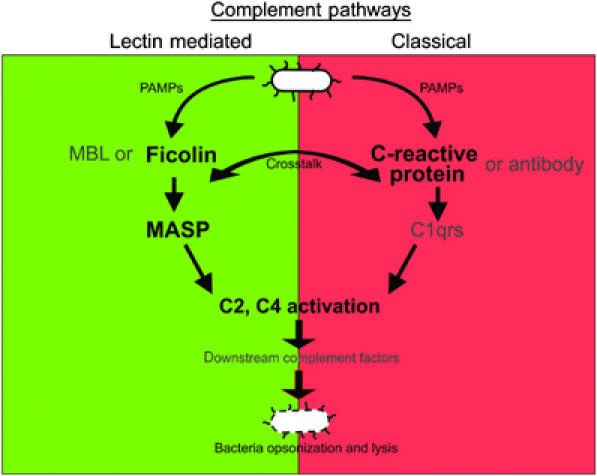
Although human C-reactive protein (CRP) becomes upregulated during septicemia, its role remains unclear, since purified CRP showed no binding to many common pathogens. Contrary to previous findings, we show that purified human CRP (hCRP) binds to Salmonella enterica, and that binding is enhanced in the presence of plasma factors. In the horseshoe crab, Carcinoscorpius rotundicauda, CRP is a major hemolymph protein. Incubation of hemolymph with a range of bacteria resulted in CRP binding to all the bacteria tested. Lipopolysaccharide-affinity chromatography of the hemolymph co-purified CRP, galactose-binding protein (GBP) and carcinolectin-5 (CL5). Yeast two-hybrid and pull-down assays suggested that these pattern recognition receptors (PRRs) form pathogen recognition complexes. We show the conservation of PRR crosstalk in humans, whereby hCRP interacts with ficolin (CL5 homologue). This interaction stabilizes CRP binding to bacteria and activates the lectin-mediated complement pathway. We propose that CRP does not act alone but collaborates with other plasma PRRs to form stable pathogen recognition complexes when targeting a wide range of bacteria for destruction.
Figures
Similar articles
-
A female-specific pentraxin, CrOctin, bridges pattern recognition receptors to bacterial phosphoethanolamine.Eur J Immunol. 2007 Dec;37(12):3477-88. doi: 10.1002/eji.200737078. Eur J Immunol. 2007. PMID: 17979155
-
Molecular interfaces of the galactose-binding protein Tectonin domains in host-pathogen interaction.J Biol Chem. 2010 Mar 26;285(13):9898-9907. doi: 10.1074/jbc.M109.059774. Epub 2010 Jan 29. J Biol Chem. 2010. PMID: 20118243 Free PMC article.
-
The macromolecular assembly of pathogen-recognition receptors is impelled by serine proteases, via their complement control protein modules.J Mol Biol. 2008 Mar 28;377(3):902-13. doi: 10.1016/j.jmb.2008.01.045. Epub 2008 Jan 30. J Mol Biol. 2008. PMID: 18279891
-
The antimicrobial properties of C-reactive protein (CRP).J Endotoxin Res. 2005;11(4):249-56. doi: 10.1179/096805105X37402. J Endotoxin Res. 2005. PMID: 16176663 Review.
-
Role of lectins in the innate immunity of horseshoe crab.Dev Comp Immunol. 1999 Jun-Jul;23(4-5):391-400. doi: 10.1016/s0145-305x(99)00019-1. Dev Comp Immunol. 1999. PMID: 10426430 Review.
Cited by
-
Elucidating the function of an ancient NF-kappaB p100 homologue, CrRelish, in antibacterial defense.Infect Immun. 2008 Feb;76(2):664-70. doi: 10.1128/IAI.00948-07. Epub 2007 Nov 26. Infect Immun. 2008. PMID: 18039837 Free PMC article.
-
The "sweet" side of a long pentraxin: how glycosylation affects PTX3 functions in innate immunity and inflammation.Front Immunol. 2013 Jan 7;3:407. doi: 10.3389/fimmu.2012.00407. eCollection 2012. Front Immunol. 2013. PMID: 23316195 Free PMC article.
-
Microbe-specific C3b deposition in the horseshoe crab complement system in a C2/factor B-dependent or -independent manner.PLoS One. 2012;7(5):e36783. doi: 10.1371/journal.pone.0036783. Epub 2012 May 7. PLoS One. 2012. PMID: 22611464 Free PMC article.
-
Preliminary evidence for obesity-associated insulin resistance in adolescents without elevations of inflammatory cytokines.Diabetol Metab Syndr. 2012 Jun 8;4(1):26. doi: 10.1186/1758-5996-4-26. Diabetol Metab Syndr. 2012. PMID: 22682228 Free PMC article.
-
Treatment of Pneumococcal Infection by Using Engineered Human C-Reactive Protein in a Mouse Model.Front Immunol. 2020 Oct 7;11:586669. doi: 10.3389/fimmu.2020.586669. eCollection 2020. Front Immunol. 2020. PMID: 33117400 Free PMC article.
References
-
- Akira S, Takeda K, Kaisho T (2001) Toll-like receptors: critical proteins linking innate and acquired immunity. Nat Immunol 2: 675–680 - PubMed
-
- Armstrong PB, Swarnakar S, Srimal S, Misquith S, Hahn EA, Aimes RT, Quigley JP (1996) A cytolytic function for a sialic acid-binding lectin that is a member of the pentraxin family of proteins. J Biol Chem 271: 14717–14721 - PubMed
-
- Black S, Kushner I, Samols D (2004) C-reactive protein. J Biol Chem 279: 48487–48490 - PubMed
-
- Blackwell JM, Searle S, Goswami T, Miller EN (2000) Understanding the multiple functions of Nramp1. Microbes Infect 2: 317–321 - PubMed
-
- Chen SC, Yen CH, Yeh MS, Huang CJ, Liu TY (2001) Biochemical properties and cDNa cloning of two new lectins from the plasma of Tachypleus tridentatus: Tachypleus plasma lectin 1 and 2+. J Biol Chem 276: 9631–9639 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
