

The FANCJ/MutLalpha interaction is required for correction of the cross-link response in FA-J cells
- PMID: 17581638
- PMCID: PMC1914102
- DOI: 10.1038/sj.emboj.7601754
The FANCJ/MutLalpha interaction is required for correction of the cross-link response in FA-J cells
Abstract
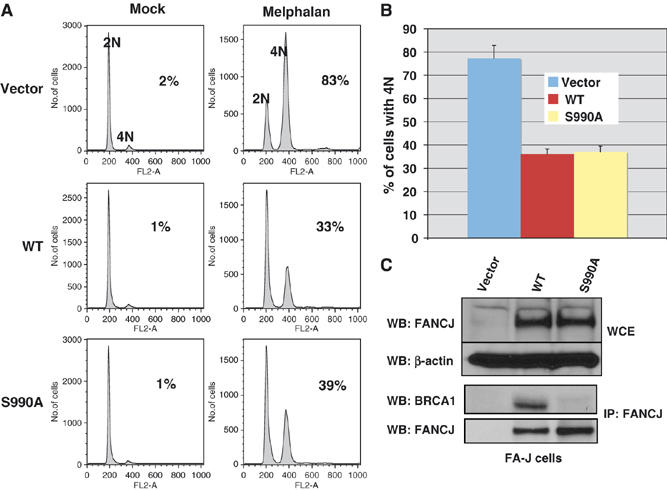
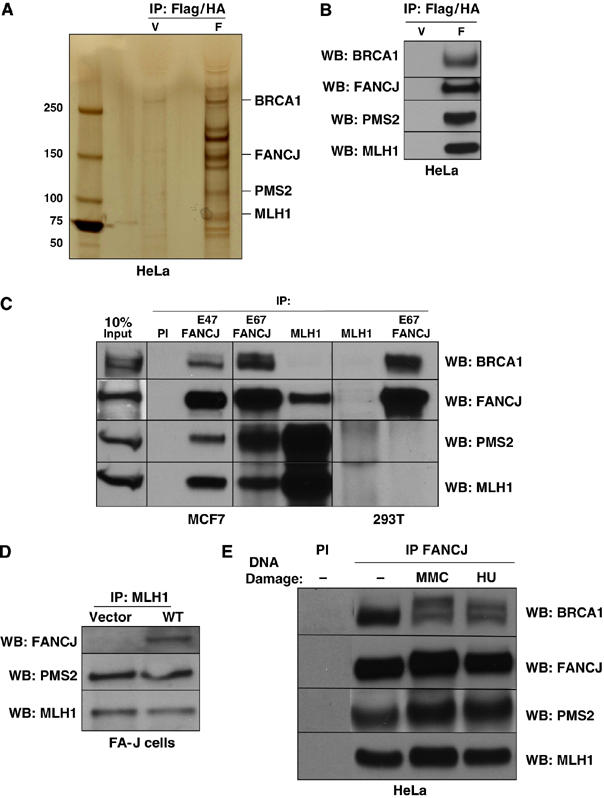
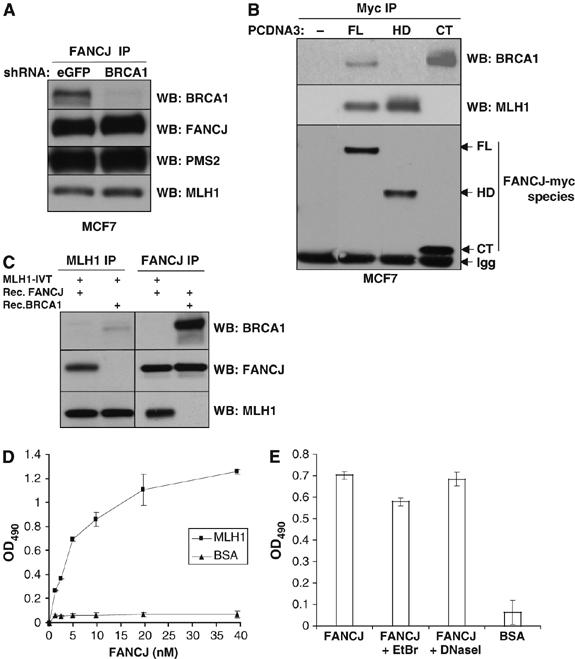
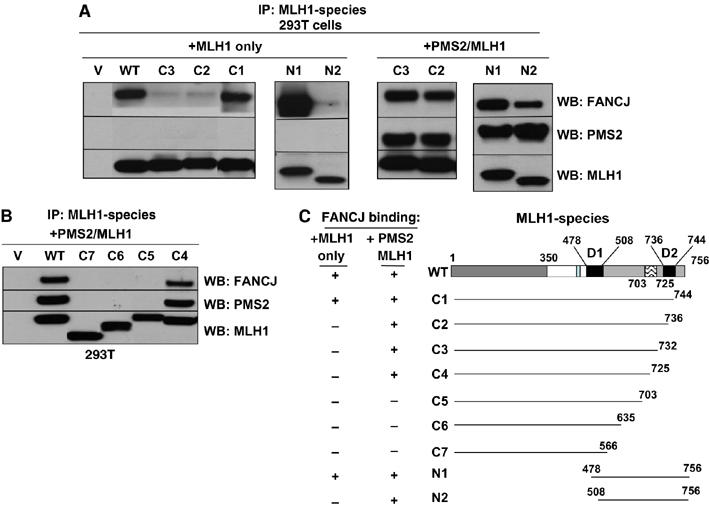
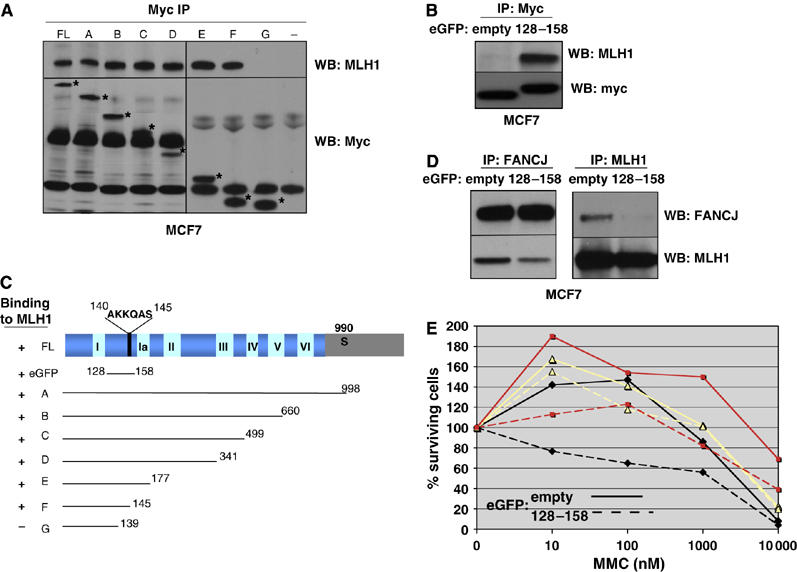
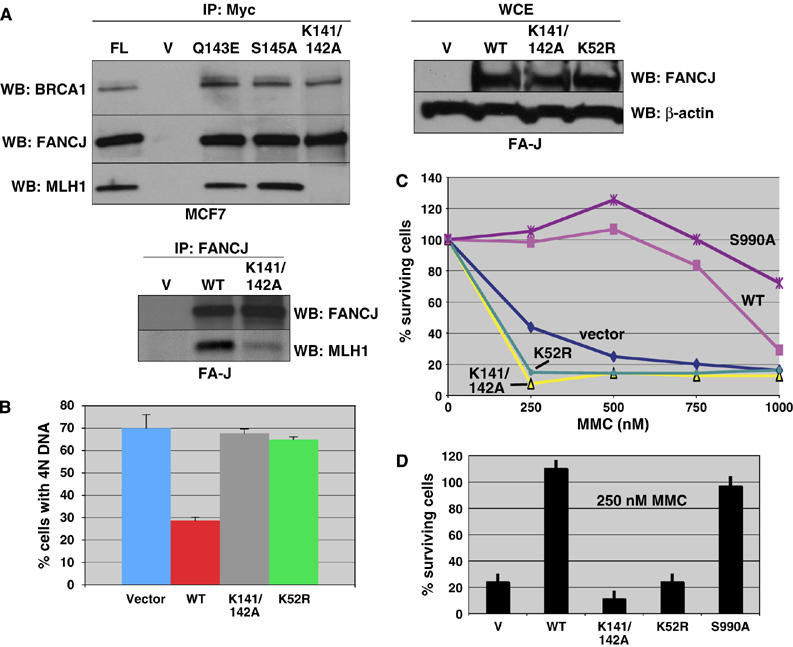
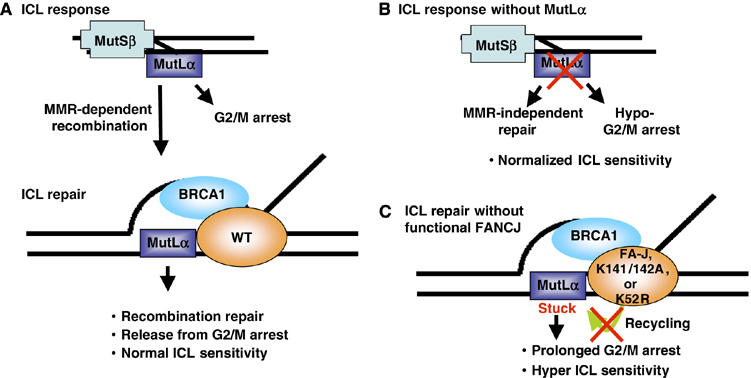
FANCJ also called BACH1/BRIP1 was first linked to hereditary breast cancer through its direct interaction with BRCA1. FANCJ was also recently identified as a Fanconi anemia (FA) gene product, establishing FANCJ as an essential tumor suppressor. Similar to other FA cells, FANCJ-null (FA-J) cells accumulate 4N DNA content in response to DNA interstrand crosslinks (ICLs). This accumulation is corrected by reintroduction of wild-type FANCJ. Here, we show that FANCJ interacts with the mismatch repair complex MutLalpha, composed of PMS2 and MLH1. Specifically, FANCJ directly interacts with MLH1 independent of BRCA1, through its helicase domain. Genetic studies reveal that FANCJ helicase activity and MLH1 binding, but not BRCA1 binding, are essential to correct the FA-J cells' ICL-induced 4N DNA accumulation and sensitivity to ICLs. These results suggest that the FANCJ/MutLalpha interaction, but not FANCJ/BRCA1 interaction, is essential for establishment of a normal ICL-induced response. The functional role of the FANCJ/MutLalpha complex demonstrates a novel link between FA and MMR, and predicts a broader role for FANCJ in DNA damage signaling independent of BRCA1.
Figures
References
-
- Akkari YM, Bateman RL, Reifsteck CA, D'Andrea AD, Olson SB, Grompe M (2001) The 4N cell cycle delay in Fanconi anemia reflects growth arrest in late S phase. Mol Genet Metab 74: 403–412 - PubMed
-
- Aquilina G, Ceccotti S, Martinelli S, Hampson R, Bignami M (1998) N-(2-chloroethyl)-N′-cyclohexyl-N-nitrosourea sensitivity in mismatch repair-defective human cells. Cancer Res 58: 135–141 - PubMed
-
- Bridge WL, Vandenberg CJ, Franklin RJ, Hiom K (2005) The BRIP1 helicase functions independently of BRCA1 in the Fanconi anemia pathway for DNA crosslink repair. Nat Genet 37: 953–957 - PubMed
-
- Brown KD, Rathi A, Kamath R, Beardsley DI, Zhan Q, Mannino JL, Baskaran R (2003) The mismatch repair system is required for S-phase checkpoint activation. Nat Genet 33: 80–84 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
