

Natural merodiploidy of the lux-rib operon of Photobacterium leiognathi from coastal waters of Honshu, Japan
- PMID: 17586644
- PMCID: PMC1951928
- DOI: 10.1128/JB.00672-07
Natural merodiploidy of the lux-rib operon of Photobacterium leiognathi from coastal waters of Honshu, Japan
Abstract
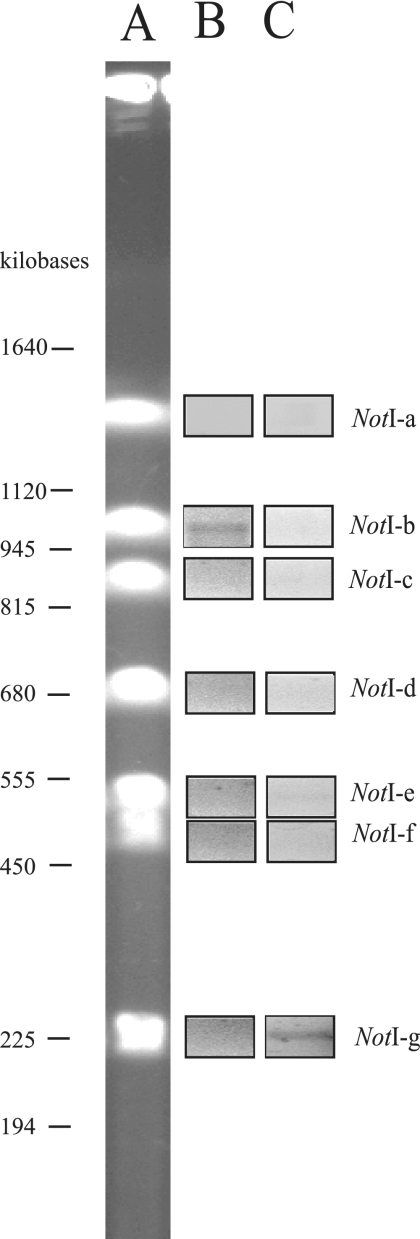
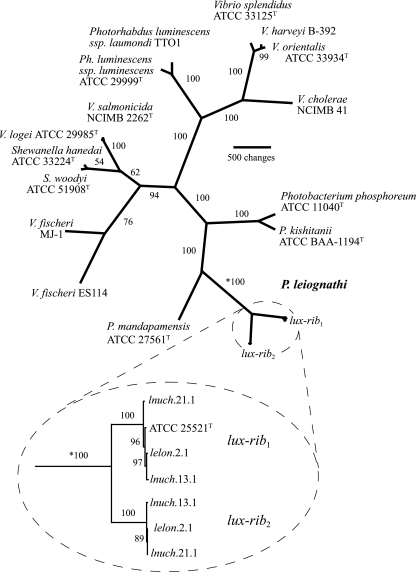
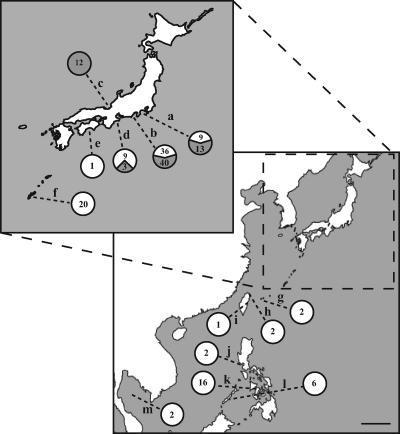
Sequence analysis of the bacterial luminescence (lux) genes has proven effective in helping resolve evolutionary relationships among luminous bacteria. Phylogenetic analysis using lux genes, however, is based on the assumptions that the lux genes are present as single copies on the bacterial chromosome and are vertically inherited. We report here that certain strains of Photobacterium leiognathi carry multiple phylogenetically distinct copies of the entire operon that codes for luminescence and riboflavin synthesis genes, luxCDABEG-ribEBHA. Merodiploid lux-rib strains of P. leiognathi were detected during sequence analysis of luxA. To define the gene content, organization, and sequence of each lux-rib operon, we constructed a fosmid library of genomic DNA from a representative merodiploid strain, lnuch.13.1. Sequence analysis of fosmid clones and genomic analysis of lnuch.13.1 defined two complete, physically separate, and apparently functional operons, designated lux-rib1 and lux-rib2. P. leiognathi strains lelon.2.1 and lnuch.21.1 were also found to carry lux-rib1 and lux-rib2, whereas ATCC 25521T apparently carries only lux-rib1. In lnuch.13.1, lelon.2.1, lnuch.21.1, and ATCC 25521T, lux-rib1 is flanked upstream by lumQ and putA and downstream by a gene for a hypothetical multidrug efflux pump. In contrast, transposase genes flank lux-rib2 of lnuch.13.1, and the chromosomal location of lux-rib2 apparently differs in lnuch.13.1, lelon.2.1, and lnuch.21.1. Phylogenetic analysis demonstrated that lux-rib1 and lux-rib2 are more closely related to each other than either one is to the lux and rib genes of other bacterial species, which rules out interspecies lateral gene transfer as the origin of lux-rib2 in P. leiognathi; lux-rib2 apparently arose within a previously unsampled or extinct P. leiognathi lineage. Analysis of 170 additional strains of P. leiognathi, for a total of 174 strains examined from coastal waters of Japan, Taiwan, the Philippine Islands, and Thailand, identified 106 strains that carry only a single lux-rib operon and 68 that carry multiple lux-rib operons. Strains bearing a single lux-rib operon were obtained throughout the geographic sampling range, whereas lux-rib merodiploid strains were found only in coastal waters of central Honshu. This is the first report of merodiploidy of lux or rib genes in a luminous bacterium and the first indication that a natural merodiploid state in bacteria can correlate with geography.
Figures


References
-
- Anderson, R. P., and J. R. Roth. 1977. Tandem genetic duplications in phage and bacteria. Annu. Rev. Microbiol. 31:473-505. - PubMed
-
- Ast, J. C., and P. V. Dunlap. 2004. Phylogenetic analysis of the lux operon distinguishes two evolutionarily distinct clades of Photobacterium leiognathi. Arch. Microbiol. 181:352-361. - PubMed
-
- Ast, J. C., and P. V. Dunlap. 2005. Phylogenetic resolution and habitat specificity of members of the Photobacterium phosphoreum species group. Environ. Microbiol. 7:1641-1654. - PubMed
-
- Ast, J. C., I. Cleenwerck, K. Engelbeen, H. Urbanczyk, F. L. Thompson, P. De Vos, and P. V. Dunlap. Photobacterium kishitanii sp. nov., a luminous marine bacterium symbiotic with deep-sea fishes. Int. J. Syst. Evol. Microbiol., in press. - PubMed
-
- Balakrishnan, S. V., and N. Langerman. 1977. The isolation of a bacterial glycoprotein with luciferase activity. Arch. Biochem. Biophys. 181:680-682. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
