

Conservation of PEX19-binding motifs required for protein targeting to mammalian peroxisomal and trypanosome glycosomal membranes
- PMID: 17586720
- PMCID: PMC1951143
- DOI: 10.1128/EC.00084-07
Conservation of PEX19-binding motifs required for protein targeting to mammalian peroxisomal and trypanosome glycosomal membranes
Abstract
Glycosomes are divergent peroxisomes found in trypanosomatid protozoa, including those that cause severe human diseases throughout much of the world. While peroxisomes are dispensable for both yeast (Saccharomyces cerevisiae and others) and mammalian cells in vitro, glycosomes are essential for trypanosomes and hence are viewed as a potential drug target. The import of proteins into the matrix of peroxisomes utilizes multiple peroxisomal membrane proteins which require the peroxin PEX19 for insertion into the peroxisomal membrane. In this report, we show that the specificity of peroxisomal membrane protein binding for Trypanosoma brucei PEX19 is very similar to those previously identified for human and yeast PEX19. Our studies show that trafficking is conserved across these distant phyla and that both a PEX19 binding site and a transmembrane domain are required for the insertion of two test proteins into the glycosomal membrane. However, in contrast to T. brucei PEX10 and PEX12, T. brucei PEX14 does not traffic to human peroxisomes, indicating that it is not recognized by the human PEX14 import mechanism.
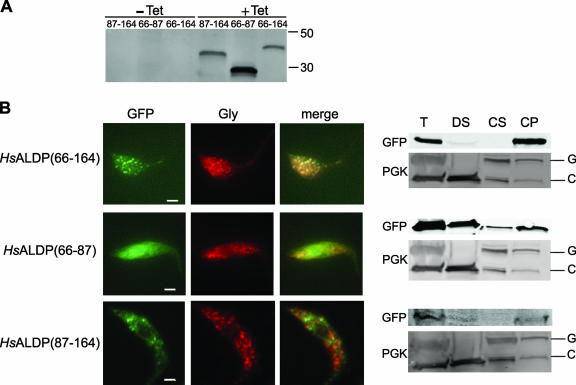
Figures





Similar articles
-
Trypanosoma brucei glycosomal ABC transporters: identification and membrane targeting.Mol Membr Biol. 2006 Mar-Apr;23(2):157-72. doi: 10.1080/09687860500460124. Mol Membr Biol. 2006. PMID: 16754359
-
Trypanosoma brucei Pex13.2 Is an Accessory Peroxin That Functions in the Import of Peroxisome Targeting Sequence Type 2 Proteins and Localizes to Subdomains of the Glycosome.mSphere. 2020 Feb 19;5(1):e00744-19. doi: 10.1128/mSphere.00744-19. mSphere. 2020. PMID: 32075879 Free PMC article.
-
Identification of trypanosomatid PEX19: functional characterization reveals impact on cell growth and glycosome size and number.Mol Biochem Parasitol. 2005 Jul;142(1):47-55. doi: 10.1016/j.molbiopara.2005.03.008. Epub 2005 Apr 7. Mol Biochem Parasitol. 2005. PMID: 15907560
-
Glycosome biogenesis in trypanosomes and the de novo dilemma.PLoS Negl Trop Dis. 2017 Apr 20;11(4):e0005333. doi: 10.1371/journal.pntd.0005333. eCollection 2017 Apr. PLoS Negl Trop Dis. 2017. PMID: 28426655 Free PMC article. Review.
-
Targeting proteins to the glycosomes of African trypanosomes.Annu Rev Microbiol. 1994;48:105-38. doi: 10.1146/annurev.mi.48.100194.000541. Annu Rev Microbiol. 1994. PMID: 7826002 Review.
Cited by
-
Using fluorescent proteins to monitor glycosome dynamics in the African trypanosome.J Vis Exp. 2014 Aug 19;(90):e51647. doi: 10.3791/51647. J Vis Exp. 2014. PMID: 25177828 Free PMC article.
-
Molecular basis of the glycosomal targeting of PEX11 and its mislocalization to mitochondrion in trypanosomes.Front Cell Dev Biol. 2023 Aug 17;11:1213761. doi: 10.3389/fcell.2023.1213761. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37664461 Free PMC article.
-
You can go your own way: The targeting signals of trypanosomatid parasites.PLoS Pathog. 2025 Aug 1;21(8):e1013326. doi: 10.1371/journal.ppat.1013326. eCollection 2025 Aug. PLoS Pathog. 2025. PMID: 40748898 Free PMC article. No abstract available.
-
Adaptive evolution of stress response genes in parasites aligns with host niche diversity.BMC Biol. 2025 Jan 13;23(1):10. doi: 10.1186/s12915-024-02091-w. BMC Biol. 2025. PMID: 39800686 Free PMC article.
-
The Arabidopsis peroxisomal ABC transporter, comatose, complements the Saccharomyces cerevisiae pxa1 pxa2Delta mutant for metabolism of long-chain fatty acids and exhibits fatty acyl-CoA-stimulated ATPase activity.J Biol Chem. 2010 Sep 24;285(39):29892-902. doi: 10.1074/jbc.M110.151225. Epub 2010 Jul 21. J Biol Chem. 2010. PMID: 20659892 Free PMC article.
References
-
- Albert, M. A., J. R. Haanstra, V. Hannaert, J. Van Roy, F. R. Opperdoes, B. M. Bakker, and P. A. Michels. 2005. Experimental and in silico analysis of glycolytic flux control in bloodstream-form Trypanosoma brucei. J. Biol. Chem. 280:28306-28315. - PubMed
-
- Albertini, M., P. Rehling, R. Erdmann, W. Girzalsky, J. A. K. W. Kiel, M. Veenhuis, and W. H. Kunau. 1997. Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell 89:83-92. - PubMed
-
- Azevedo, J. E., J. Costa-Rodrigues, C. P. Guimaraes, M. E. Oliveira, and C. Sa-Miranda. 2004. Protein translocation across the peroxisomal membrane. Cell Biochem. Biophys. 41:451-468. - PubMed
-
- Banerjee, S. K., P. S. Kessler, T. Saveria, and M. Parsons. 2005. Identification of trypanosomatid PEX19: functional characterization reveals impact on cell growth and glycosome size and number. Mol. Biochem. Parasitol. 142:47-55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
