

Negative feedback regulation of hypophysiotropic thyrotropin-releasing hormone (TRH) synthesizing neurons: role of neuronal afferents and type 2 deiodinase
- PMID: 17588648
- PMCID: PMC2000455
- DOI: 10.1016/j.yfrne.2007.04.002
Negative feedback regulation of hypophysiotropic thyrotropin-releasing hormone (TRH) synthesizing neurons: role of neuronal afferents and type 2 deiodinase
Abstract
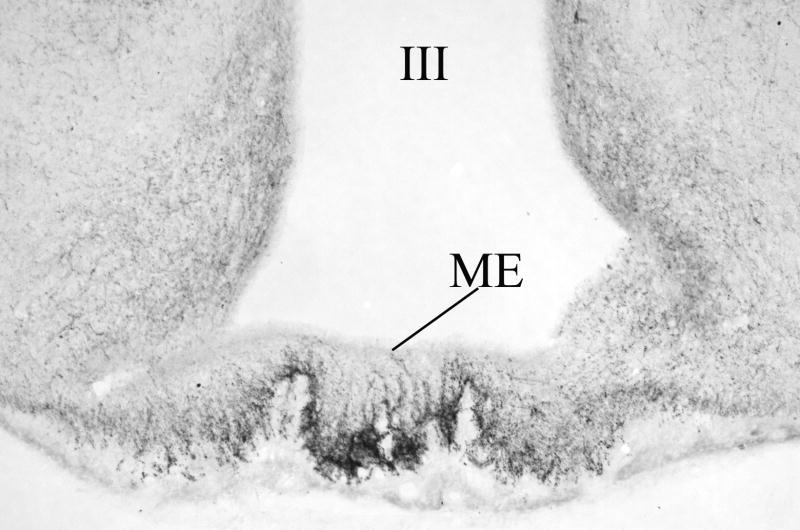
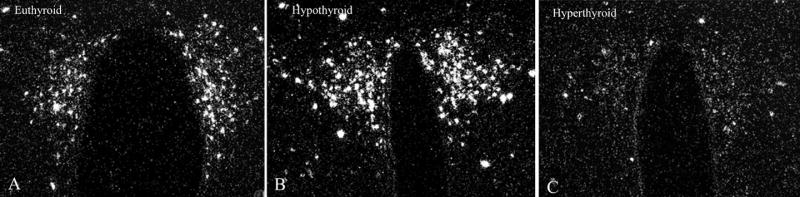
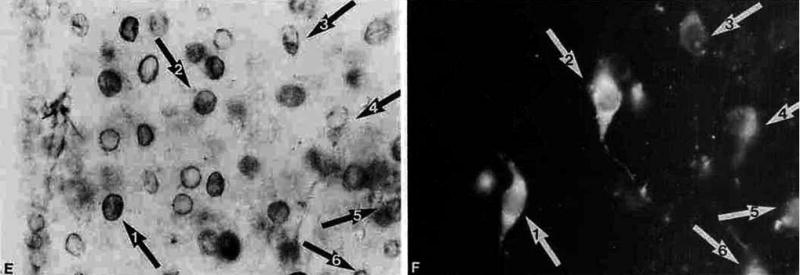
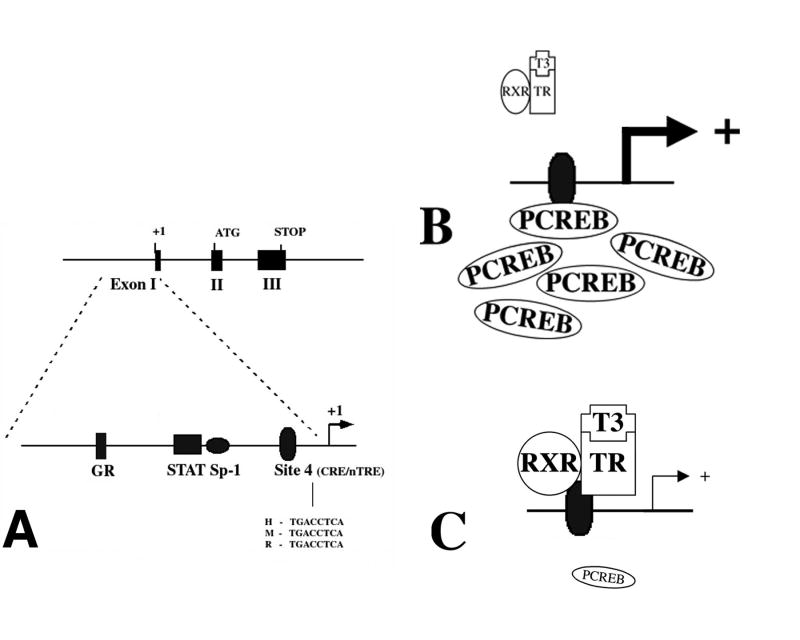
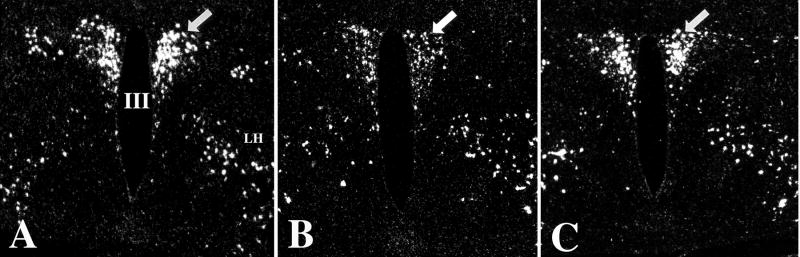
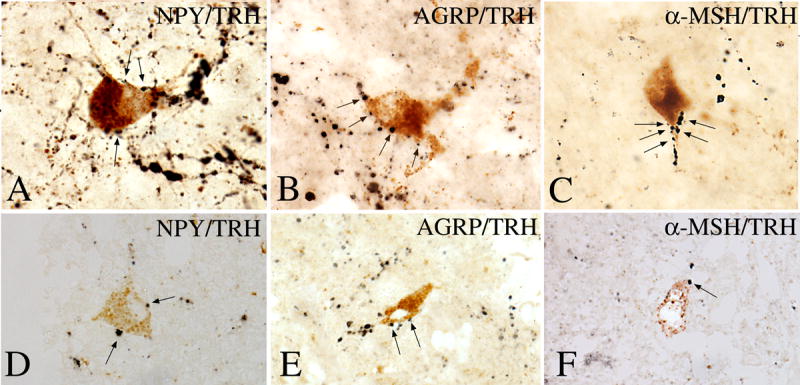
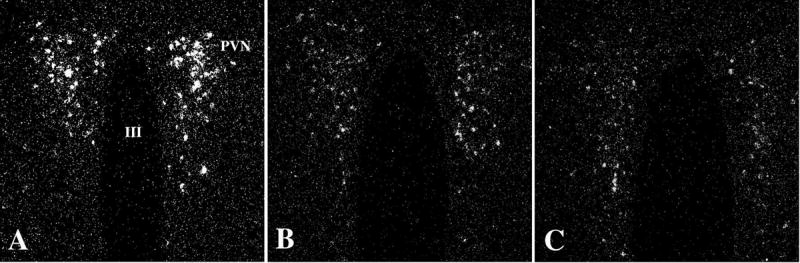
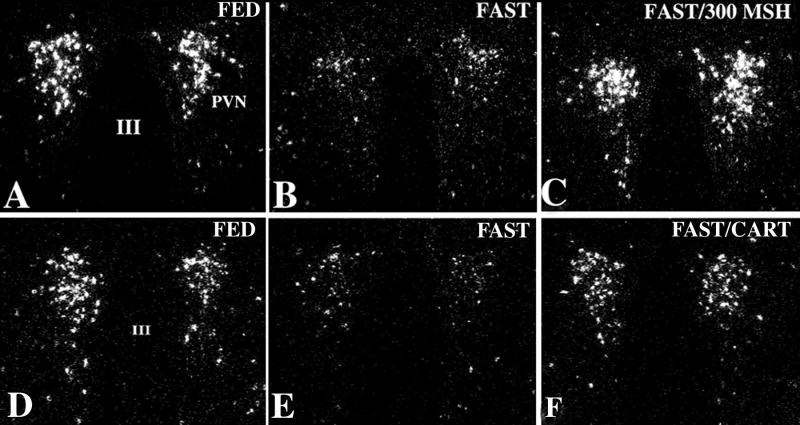
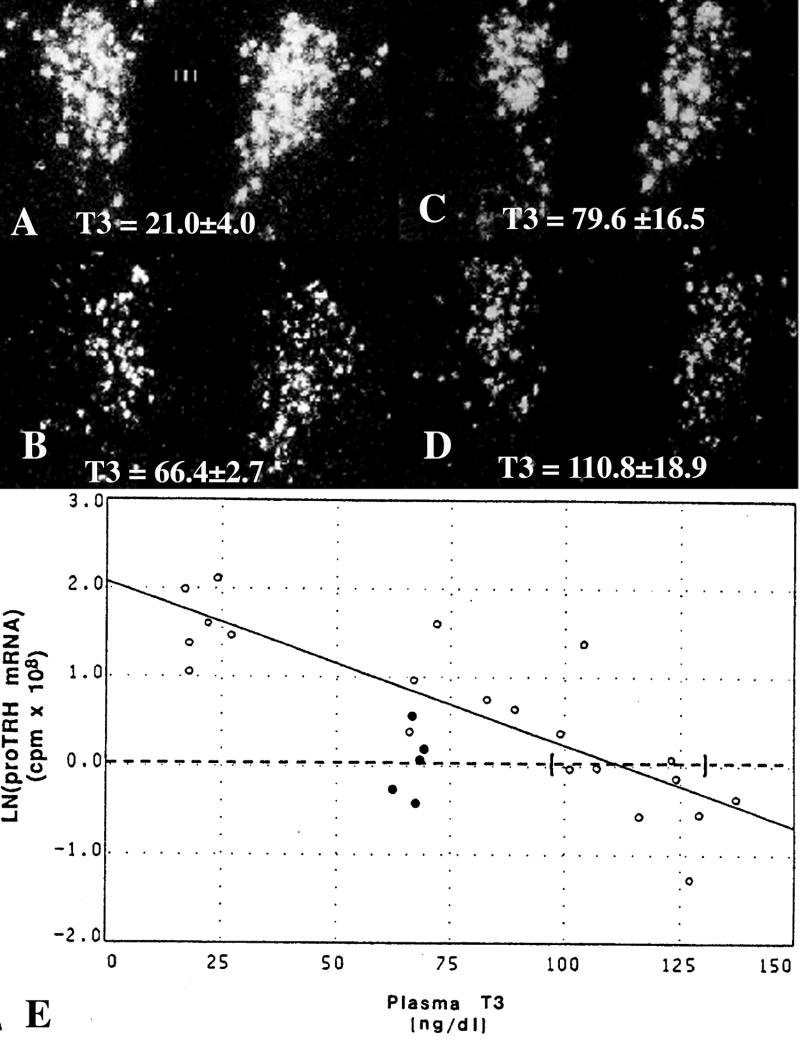
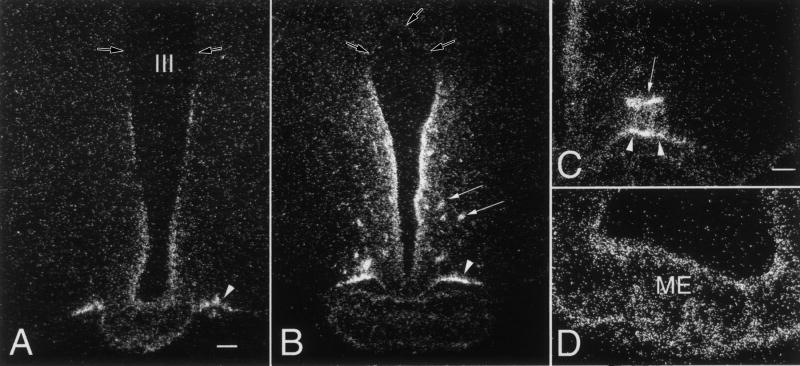
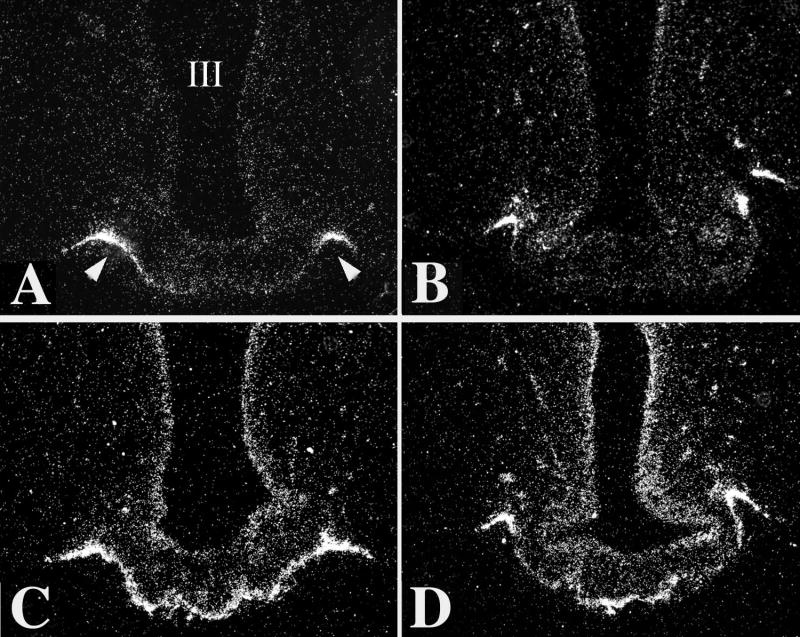
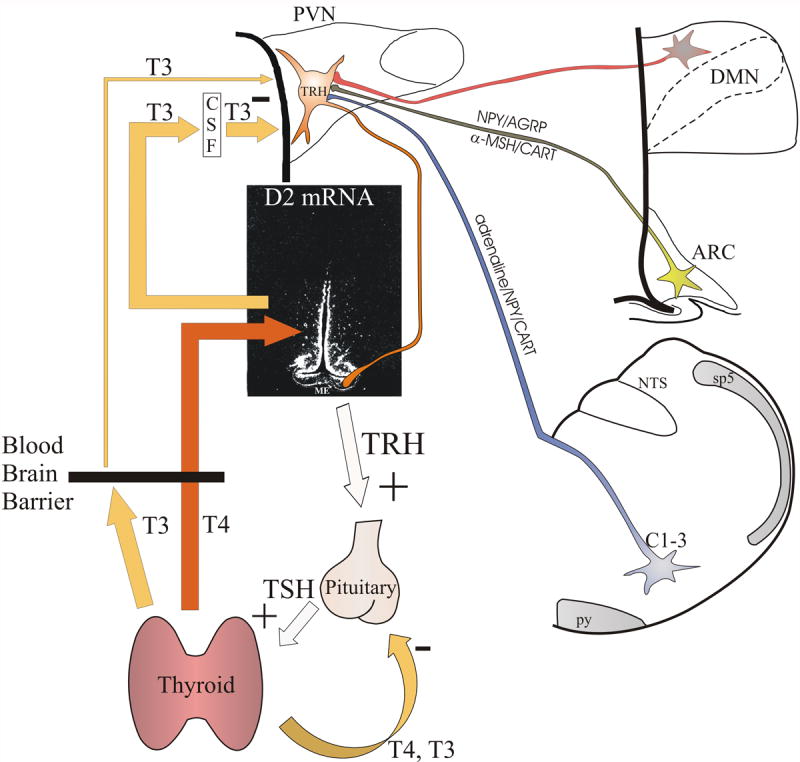
Hypophysiotropic thyrotropin-releasing hormone (TRH): synthesizing neurons reside in the hypothalamic paraventricular nucleus (PVN) and are the central regulators of the hypothalamic-pituitary-thyroid (HPT) axis. TRH synthesis and release from these neurons are primarily under negative feedback regulation by thyroid hormone. Under certain conditions such as cold exposure and fasting, however, inputs from neurons in the brainstem and hypothalamic arcuate and dorsomedial nuclei alter the set point for negative feedback through regulation of CREB phosphorylation. Thus, during cold exposure, adrenergic neurons stimulate the HPT axis, while fasting-induced central hypothyroidism is mediated through an arcuato-paraventricular pathway. Feedback regulation of TRH neurons may also be modified by local tissue levels of thyroid hormone regulated by the activation of type 2 iodothyronine deiodinase (D2), the primary enzyme in the brain that catalyzes T4 to T3 conversion. During infection, endotoxin or endotoxin induced cytokines increase D2 activity in the mediobasal hypothalamus, which by inducing local hyperthyroidism, may play an important role in infection-induced inhibition of hypophysiotropic TRH neurons.
Figures

References
-
- Lechan RM. Update on thyrotropin-releasing hormone. Thyroid Today. 1993;16:1–12.
-
- Nillni EA, Sevarino KA. The biology of prothyrotropin-releasing hormone-derived peptides. Endocr Rev. 1999;20:599–648. - PubMed
-
- Bradbury AF, Finnie MD, Smyth DG. Mechanism of C-terminal amide formation by pituitary enzymes. Nature. 1982;298:686–688. - PubMed
-
- Lechan RM, Segerson TP. Pro-TRH gene expression and precursor peptides in rat brain. Observations by hybridization analysis and immunocytochemistry. Ann N Y Acad Sci. 1989;553:29–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
