

Extracellular matrix, mechanotransduction and structural hierarchies in heart tissue engineering
- PMID: 17588874
- PMCID: PMC2440395
- DOI: 10.1098/rstb.2007.2114
Extracellular matrix, mechanotransduction and structural hierarchies in heart tissue engineering
Abstract
The spatial and temporal scales of cardiac organogenesis and pathogenesis make engineering of artificial heart tissue a daunting challenge. The temporal scales range from nanosecond conformational changes responsible for ion channel opening to fibrillation which occurs over seconds and can lead to death. Spatial scales range from nanometre pore sizes in membrane channels and gap junctions to the metre length scale of the whole cardiovascular system in a living patient. Synchrony over these scales requires a hierarchy of control mechanisms that are governed by a single common principle: integration of structure and function. To ensure that the function of ion channels and contraction of muscle cells lead to changes in heart chamber volume, an elegant choreography of metabolic, electrical and mechanical events are executed by protein networks composed of extracellular matrix, transmembrane integrin receptors and cytoskeleton which are functionally connected across all size scales. These structural control networks are mechanoresponsive, and they process mechanical and chemical signals in a massively parallel fashion, while also serving as a bidirectional circuit for information flow. This review explores how these hierarchical structural networks regulate the form and function of living cells and tissues, as well as how microfabrication techniques can be used to probe this structural control mechanism that maintains metabolic supply, electrical activation and mechanical pumping of heart muscle. Through this process, we delineate various design principles that may be useful for engineering artificial heart tissue in the future.
Figures
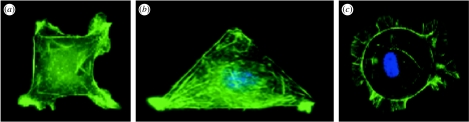
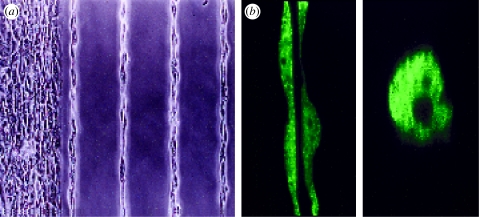
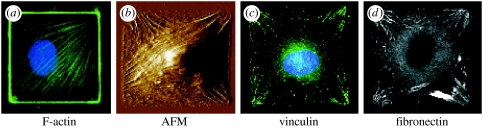


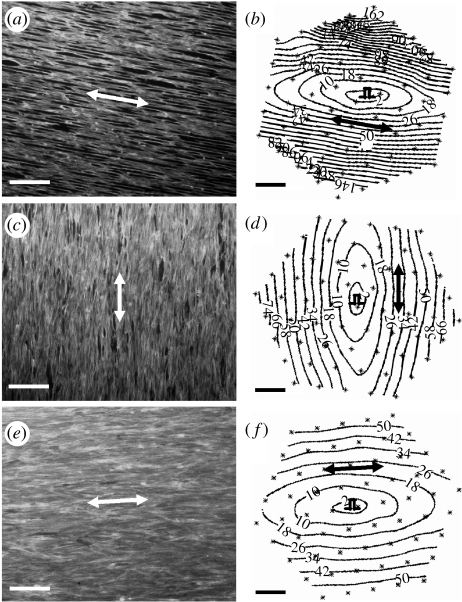
References
-
- Adler C.P, Ringlage W.P, Bohm N. DNA content and cell number in heart and liver of children. Comparable biochemical, cytophotometric and histological investigations. Pathol. Res. Pract. 1981;172:25–41. [Author's transl.] - PubMed
-
- Alenghat F.J, Ingber D.E. Mechanotransduction: all signals point to cytoskeleton, matrix, and integrins. Sci. STKE. 2002;2002:PE6. - PubMed
-
- Badylak S, Obermiller J, Geddes L, Matheny R. Extracellular matrix for myocardial repair. Heart Surg. Forum. 2003;6:E20–E26. - PubMed
-
- Baicu C.F, Stroud J.D, Livesay V.A, Hapke E, Holder J, Spinale F.G, Zile M.R. Changes in extracellular collagen matrix alter myocardial systolic performance. Am. J. Physiol. Heart Circ. Physiol. 2003;284:H122–H132. - PubMed
-
- Bick R.J, Snuggs M.B, Poindexter B.J, Buja L.M, Van Winkle W.B. Physical, contractile and calcium handling properties of neonatal cardiac myocytes cultured on different matrices. Cell Adhes. Commun. 1998;6:301–310. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
