

Aromatase excess in cancers of breast, endometrium and ovary
- PMID: 17590327
- PMCID: PMC2766613
- DOI: 10.1016/j.jsbmb.2007.05.027
Aromatase excess in cancers of breast, endometrium and ovary
Abstract
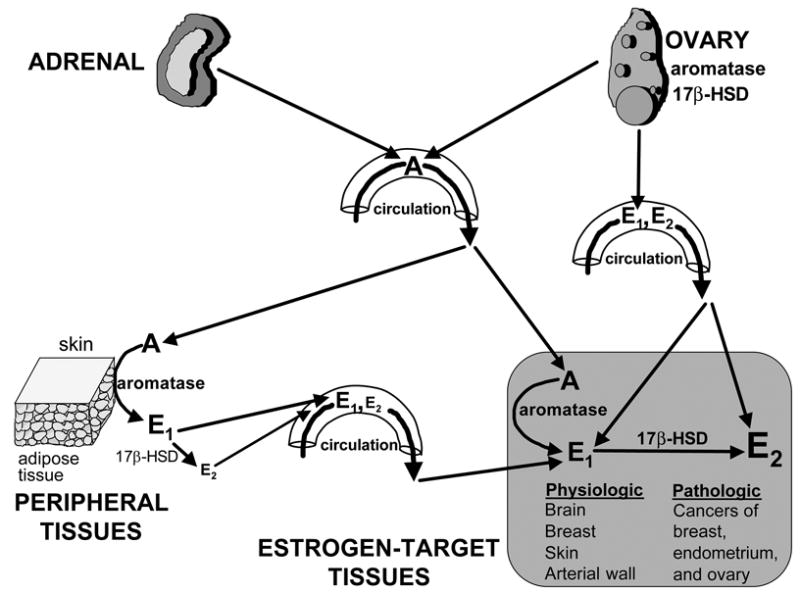
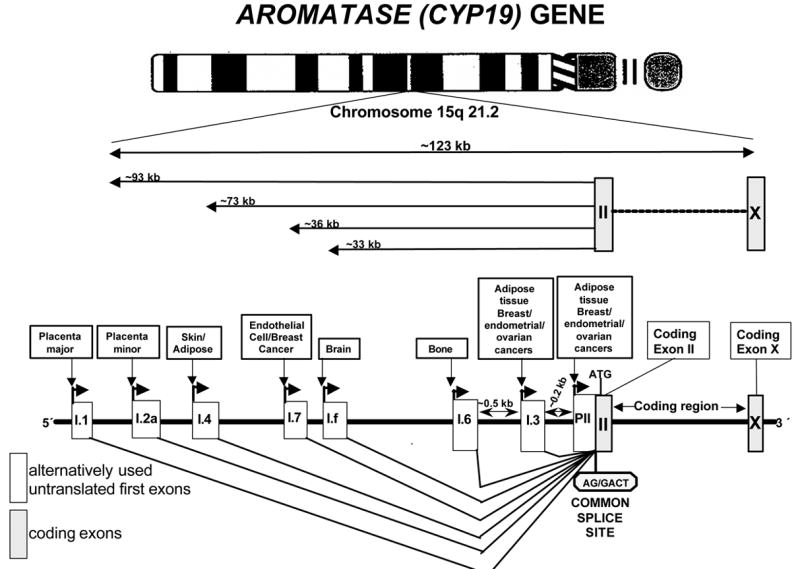
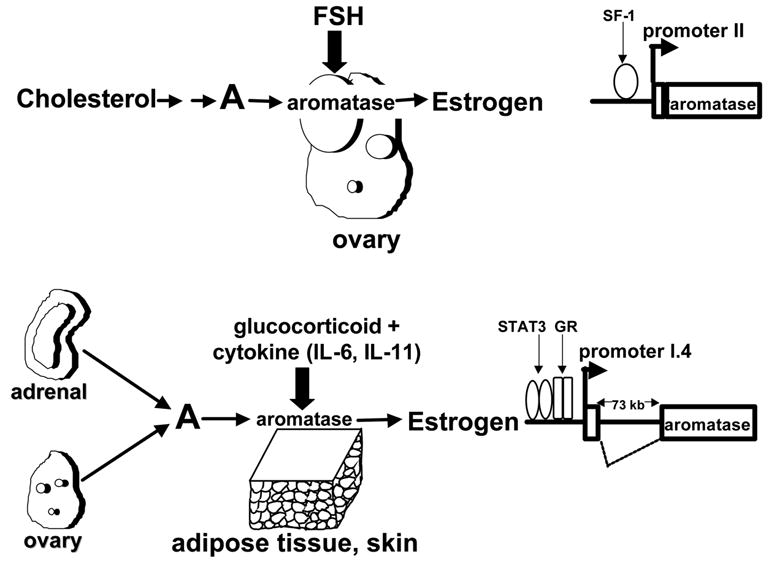
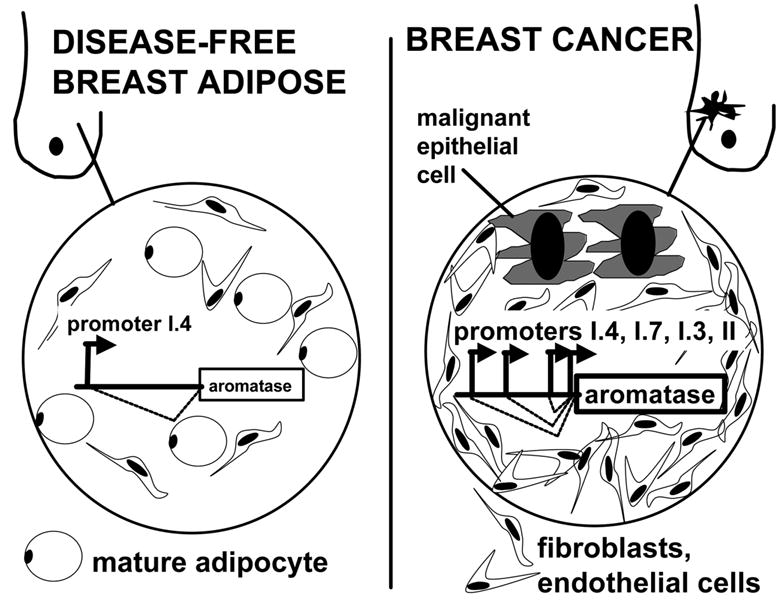
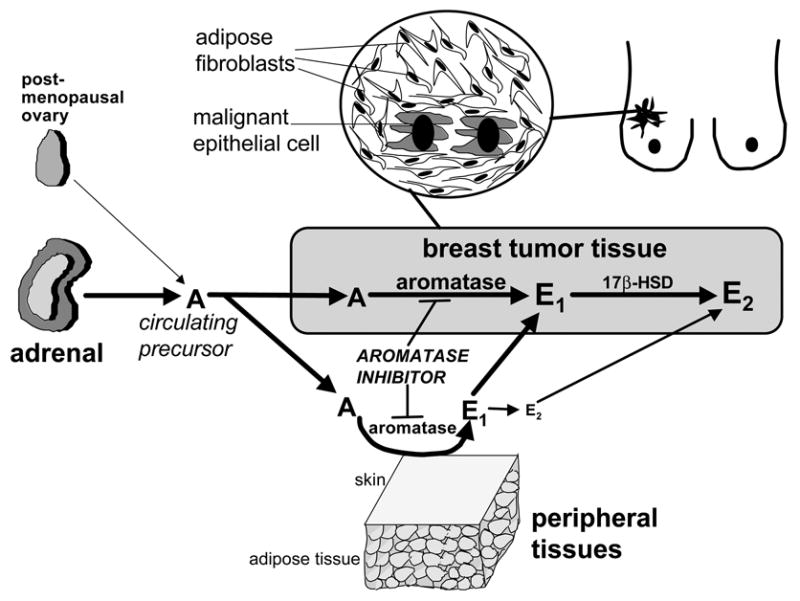
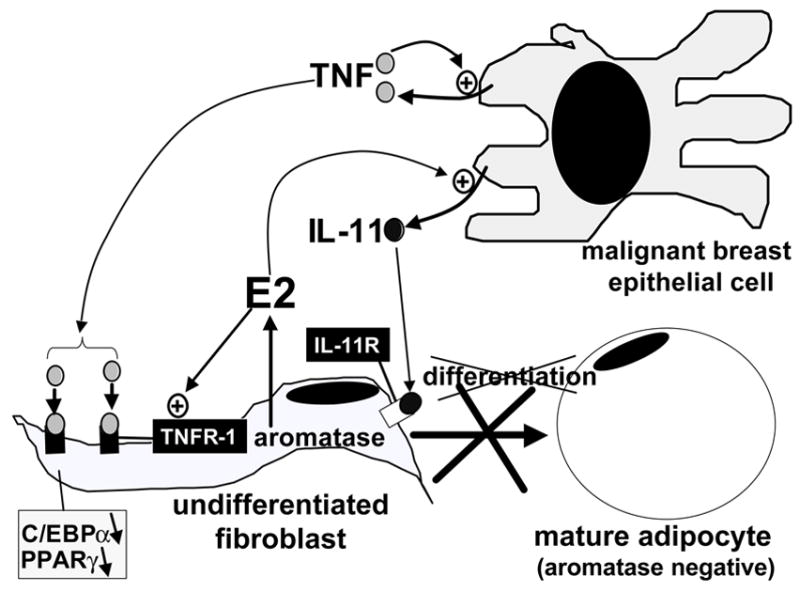
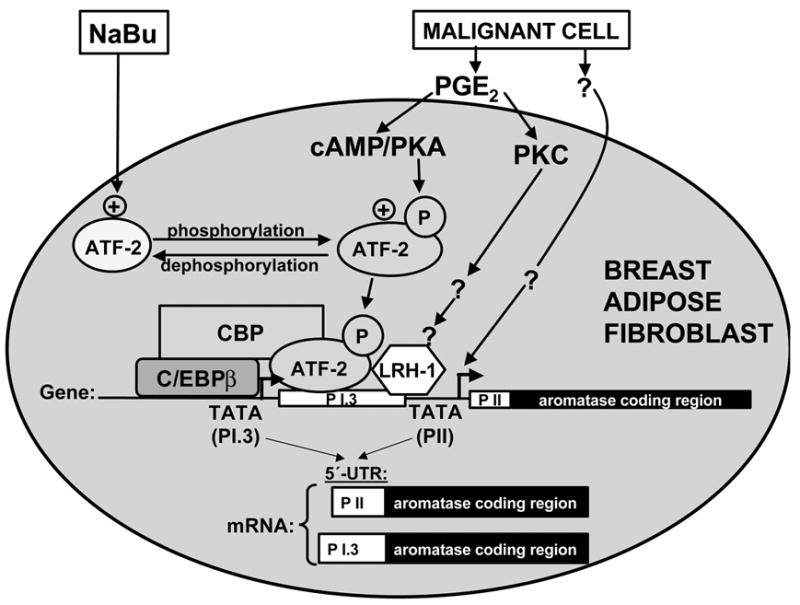
Pathogenesis and growth of three common women's cancers (breast, endometrium and ovary) are linked to estrogen. A single gene encodes the key enzyme for estrogen biosynthesis named aromatase, inhibition of which effectively eliminates estrogen production in the entire body. Aromatase inhibitors successfully treat breast cancer, whereas their roles in endometrial and ovarian cancers are less clear. Ovary, testis, adipose tissue, skin, hypothalamus and placenta express aromatase normally, whereas breast, endometrial and ovarian cancers overexpress aromatase and produce local estrogen exerting paracrine and intracrine effects. Tissue-specific promoters distributed over a 93-kb regulatory region upstream of a common coding region alternatively control aromatase expression. A distinct set of transcription factors regulates each promoter in a signaling pathway- and tissue-specific manner. In cancers of breast, endometrium and ovary, aromatase expression is primarly regulated by increased activity of the proximally located promoter I.3/II region. Promoters I.3 and II lie 215 bp from each other and are coordinately stimulated by PGE(2) via a cAMP-PKA-dependent pathway. In breast adipose fibroblasts exposed to PGE(2) secreted by malignant epithelial cells, PKC is also activated, and this potentiates cAMP-PKA-dependent induction of aromatase. Thus, inflammatory substances such as PGE(2) may play important roles in inducing local production of estrogen that promotes tumor growth.
Figures
References
-
- Simpson ER, Clyne C, Rubin G, Boon WC, Robertson K, Britt K, Speed C, Jones M. Aromatase--a brief overview. Annu Rev Physiol. 2002;64:93–127. - PubMed
-
- Sebastian S, Bulun SE. A highly complex organization of the regulatry region of the human CYP19 (aromatase) gene revealed by the human genome project. J Clin Endocrinol Metab. 2001;86:4600–4602. - PubMed
-
- Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham-Lorence S, Amarneh B, Ito Y, Fisher CR, Michael MD, Mendelson CR, Bulun SE. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocrine Reviews. 1994;15:342–355. - PubMed
-
- Bulun SE, Simpson ER. Competitive RT-PCR analysis indicates levels of aromatase cytochrome P450 transcripts in adipose tissue of buttocks, thighs, and abdomen of women increase with advancing age. Journal of Clinical Endocrinology and Metabolism. 1994;78:428–432. - PubMed
-
- Grodin JM, Siiteri PK, MacDonald PC. Source of estrogen production in postmenopausal women. Journal of Clinical Endocrinology and Metabolism. 1973;36:207–214. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
