

Horizontal saccade disconjugacy in strabismic monkeys
- PMID: 17591880
- PMCID: PMC2562538
- DOI: 10.1167/iovs.06-0955
Horizontal saccade disconjugacy in strabismic monkeys
Abstract
Purpose: Previous studies have shown that binocular coordination during saccadic eye movement is affected in humans with large strabismus. The purpose of this study was to examine the conjugacy of saccadic eye movements in monkeys with sensory strabismus.
Methods: The authors recorded binocular eye movements in four strabismic monkeys and one unaffected monkey. Strabismus was induced by first occluding one eye for 24 hours, switching the occluder to the fellow eye for the next 24 hours, and repeating this pattern of daily alternating monocular occlusion for the first 4 to 6 months of life. Horizontal saccades were measured during monocular viewing when the animals were 2 to 3 years of age.
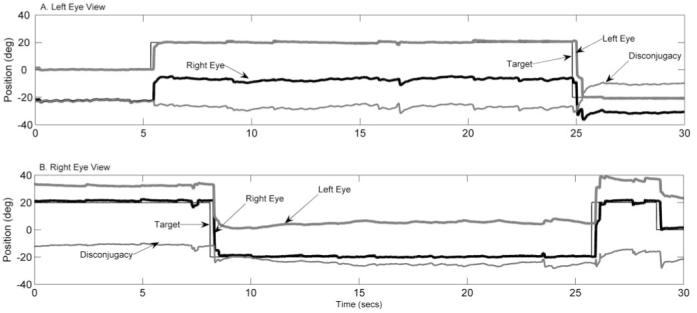
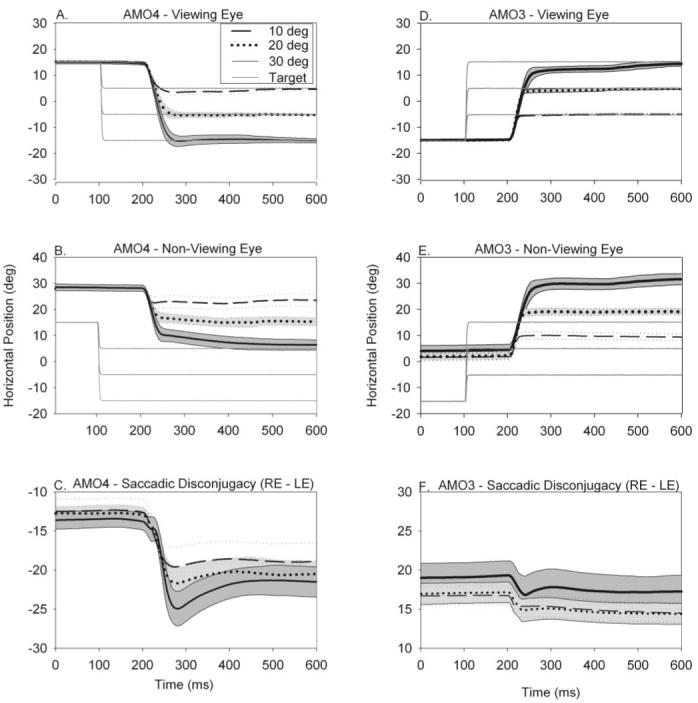
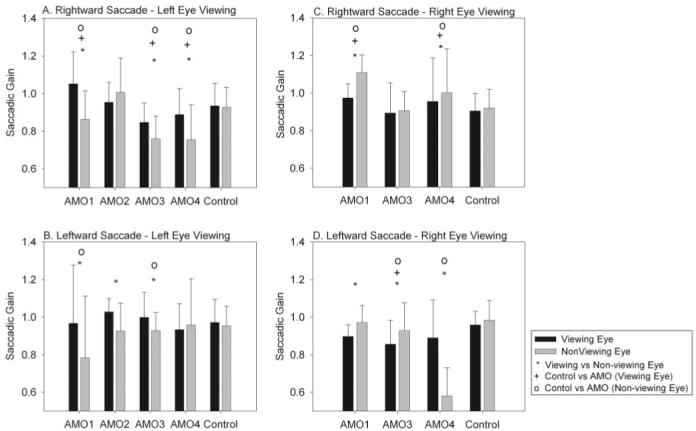
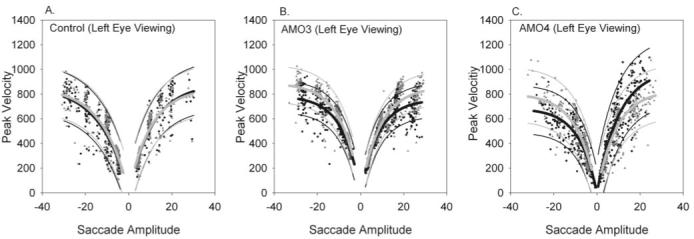
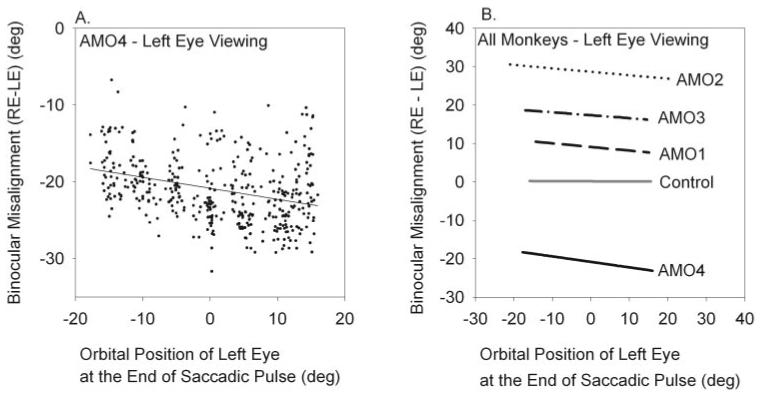
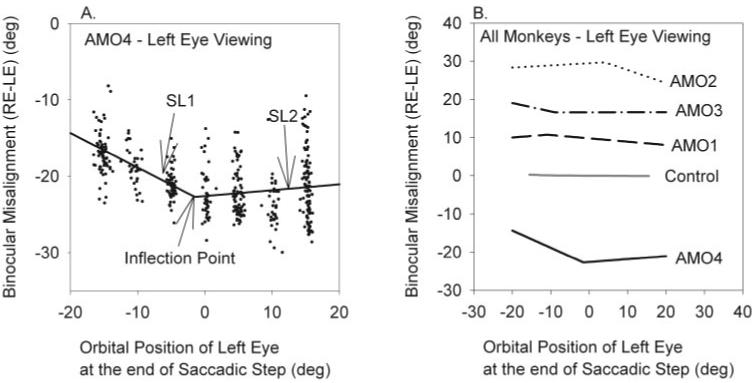
Results: Horizontal saccade testing during monocular viewing showed that the amplitude of saccades in the nonviewing eye was usually different from that in the viewing eye (saccade disconjugacy). The amount of saccade disconjugacy varied among animals as a function of the degree of ocular misalignment as measured in primary gaze. Saccade disconjugacy also increased with eccentric orbital positions of the nonviewing eye. If the saccade disconjugacy was large, there was an immediate postsaccadic drift for less than 200 ms. The control animal showed none of these effects.
Conclusions: As do humans with large strabismus, strabismic monkey display disconjugate saccadic eye movements. Saccade disconjugacy varies with orbital position and increases as a function of ocular misalignment as measured in primary gaze. This type of sensory-induced strabismus serves as a useful animal model to investigate the neural or mechanical factors responsible for saccade disconjugacy observed in humans with strabismus.
Figures
References
-
- von Noorden GK. Binocular Vision and Ocular Motility: Theory and Management of Strabismus. 5th ed. CV Mosby; St. Louis: 1996.
-
- Leigh RJ, Zee DS. The Neurology of Eye Movements. 3rd ed. Oxford University Press; New York: 1999. Contemporary Neurology Series.
-
- Boothe RG, Dobson V, Teller DY. Postnatal development of vision in human and nonhuman primates. Ann Rev Neurosci. 1985;8:495–545. - PubMed
-
- Lorenz B. Genetics of isolated and syndromic strabismus: facts and perspectives. Strabismus. 2002;10:147–156. - PubMed
-
- Maxwell GF, Lemij HG, Collewijn H. Conjugacy of saccades in deep amblyopia. Invest Ophthalmol Vis Sci. 1995;36:2514–2522. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
