

Differential factor requirement to assemble translation initiation complexes at the alternative start codons of foot-and-mouth disease virus RNA
- PMID: 17592045
- PMCID: PMC1924898
- DOI: 10.1261/rna.469707
Differential factor requirement to assemble translation initiation complexes at the alternative start codons of foot-and-mouth disease virus RNA
Abstract
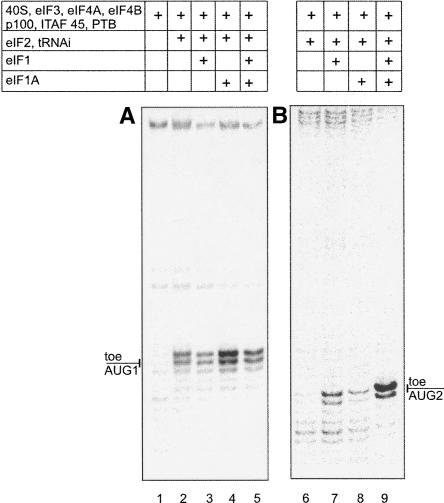
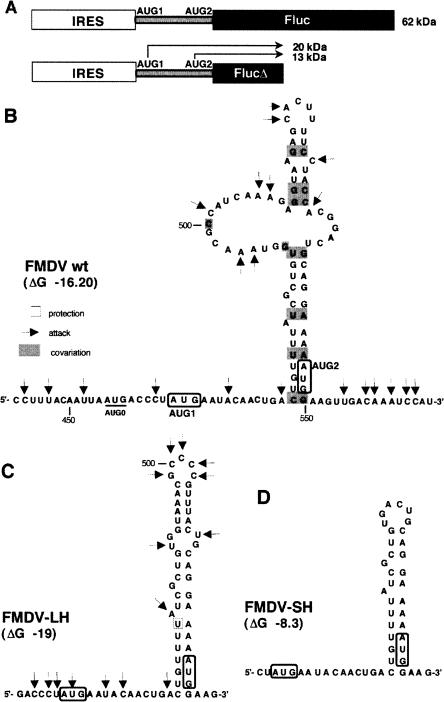
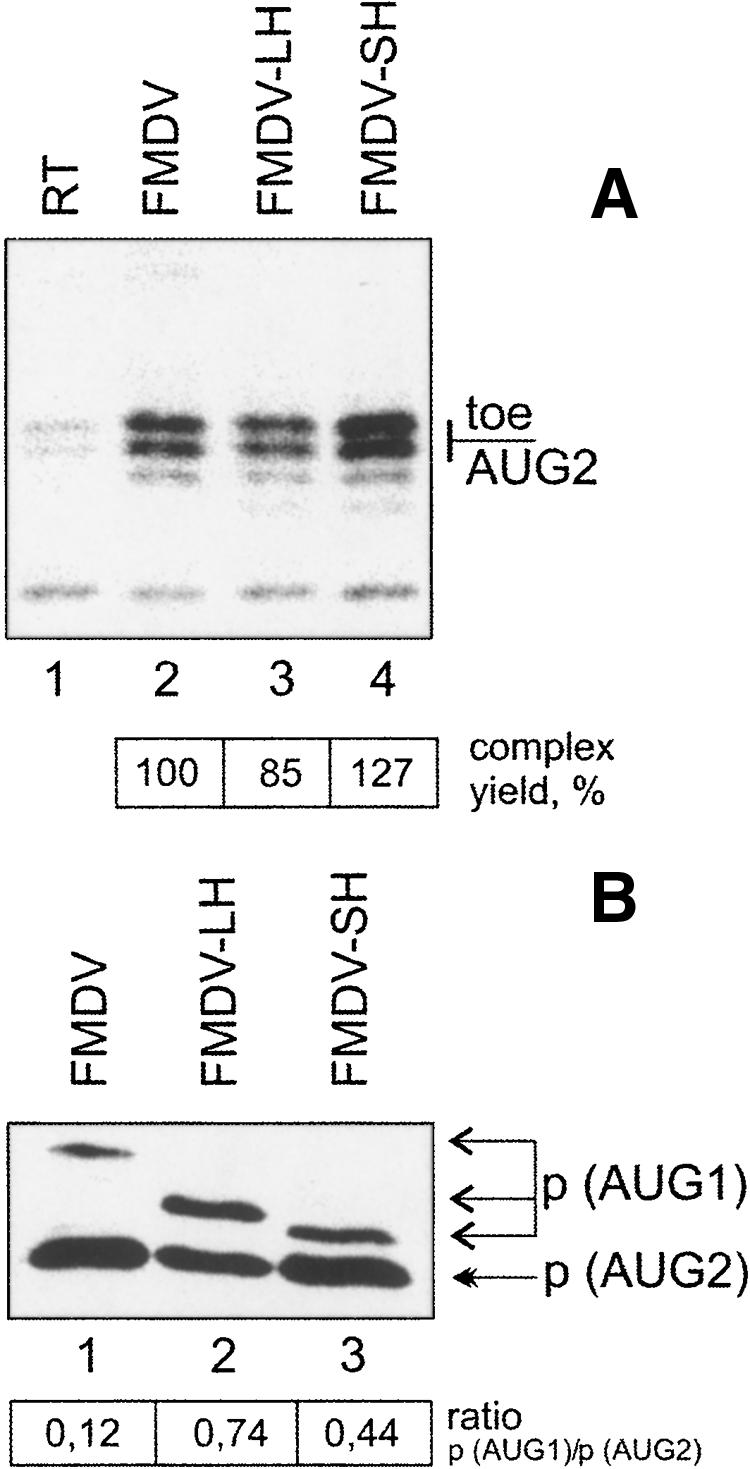
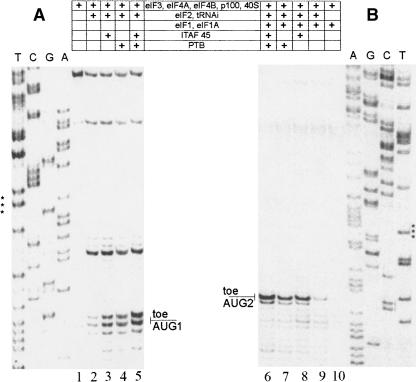
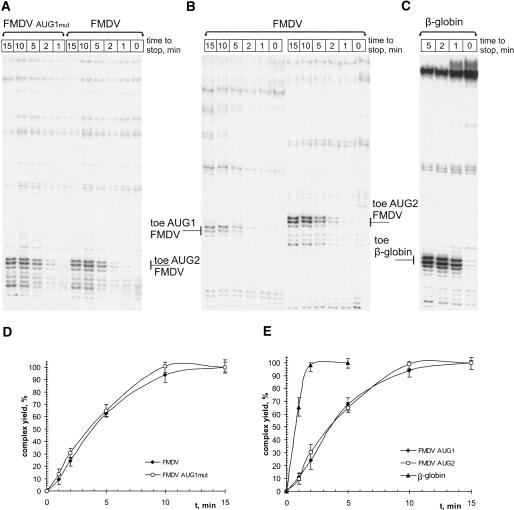
The foot-and-mouth disease virus (FMDV) RNA contains two in-frame AUG codons separated by 84 nt that direct translation initiation of the viral polyprotein. The mechanism of initiation at the IRES-proximal AUG codon (AUG1) has been previously analyzed, whereas no data on factor requirements for AUG2 have been reported. Here, using the method of 48S translation initiation complex reconstitution, we show that eIF1 is indispensable in forming the 48S initiation complex at AUG2. In contrast, it reduces the assembly of this complex at AUG1. Stabilization of a stem-loop between the initiation triplets induces a small decrease in the toeprint intensity at AUG2, accompanied by an increase in the AUG1/AUG2 ratio as well as a moderate reduction of protein synthesis initiated at AUG2 in transfected cells. PTB and ITAF45 exerted an additive positive effect on the 48S complex at AUG2, although a substantial reconstitution on both AUGs occurs on omission of either of these proteins. Relative to the beta-globin mRNA, the 48S complex formation at AUG1 and AUG2 is slow and occurs with the same kinetics as revealed by the "kinetic" toeprint assay. Mutation of AUG1 to AUA does not abrogate protein synthesis in transfected cells, and has no effect on the rate of the 48S complex formation at AUG2. We conclude that the AUG2 initiation region is selected independently of 48S complex formation at the upstream AUG1. The kinetic toeprint assay also shows that cap-dependent assembly of the 48S complex in vitro occurs faster than the FMDV IRES-mediated complex assembly.
Figures
References
-
- Belsham, G.J., Jackson, R.J. Translation initiation on picornavirus RNA. In: Sonenberg N., et al., editors. Translational control of gene expression. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 2000. pp. 869–900.
-
- Belsham, G.J., Martinez-Salas, E. Genome organization, translation, and replication of foot-and-mouth disease virus RNA. In: Domingo E., Sobrino F., editors. Foot-and-mouth disease: Current perspectives. Horizon Scientific Press; Norwich, UK: 2004. pp. 19–52.
-
- Borovjagin, A., Pestova, T., Shatsky, I. Pyrimidine tract binding protein strongly stimulates in vitro encephalomyocarditis virus RNA translation at the level of preinitiation complex formation. FEBS Lett. 1994;351:299–302. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous