

Csrp1 regulates dynamic cell movements of the mesendoderm and cardiac mesoderm through interactions with Dishevelled and Diversin
- PMID: 17592114
- PMCID: PMC2040889
- DOI: 10.1073/pnas.0702000104
Csrp1 regulates dynamic cell movements of the mesendoderm and cardiac mesoderm through interactions with Dishevelled and Diversin
Abstract
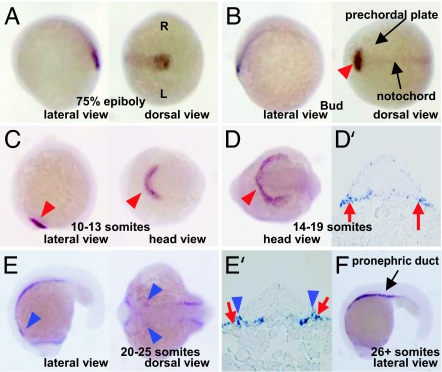
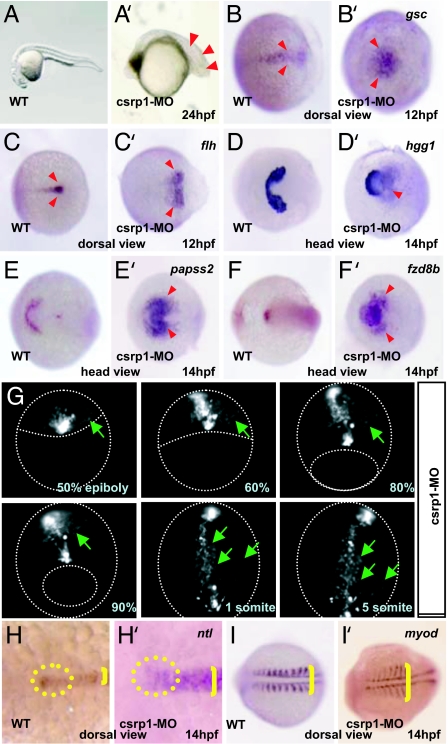
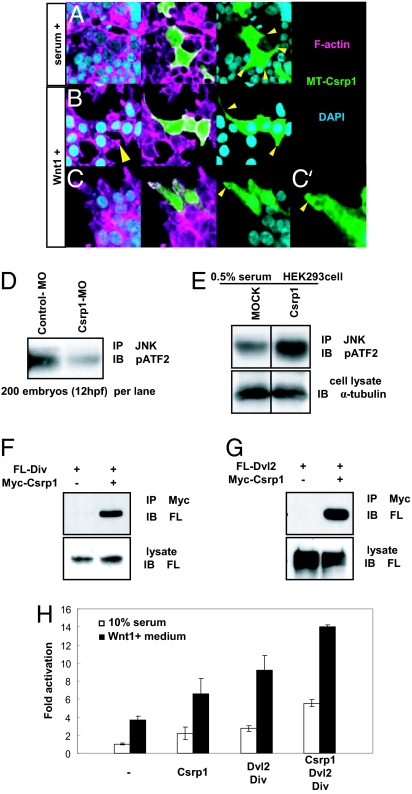
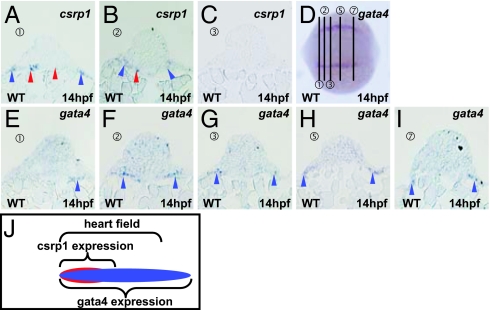
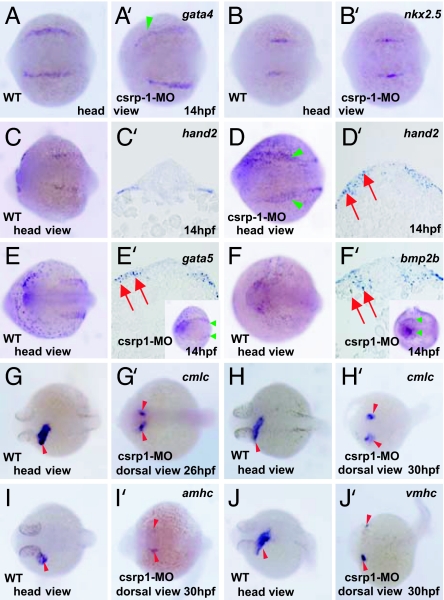
Zebrafish Csrp1 is a member of the cysteine- and glycine-rich protein (CSRP) family and is expressed in the mesendoderm and its derivatives. Csrp1 interacts with Dishevelled 2 (Dvl2) and Diversin (Div), which control cell morphology and other dynamic cell behaviors via the noncanonical Wnt and JNK pathways. When csrp1 message is knocked down, abnormal convergent extension cell movement is induced, resulting in severe deformities in midline structures. In addition, cardiac bifida is induced as a consequence of defects in cardiac mesoderm cell migration. Our data highlight Csrp1 as a key molecule of the noncanonical Wnt pathway, which orchestrates cell behaviors during dynamic morphogenetic movements of tissues and organs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Cooperation of polarized cell intercalations drives convergence and extension of presomitic mesoderm during zebrafish gastrulation.J Cell Biol. 2008 Jan 14;180(1):221-32. doi: 10.1083/jcb.200704150. J Cell Biol. 2008. PMID: 18195109 Free PMC article.
-
Diversin regulates heart formation and gastrulation movements in development.Proc Natl Acad Sci U S A. 2006 Oct 24;103(43):15900-5. doi: 10.1073/pnas.0603808103. Epub 2006 Oct 10. Proc Natl Acad Sci U S A. 2006. PMID: 17032765 Free PMC article.
-
[KLHL12/Cullin-3 is a new E3 ubiquitin ligase negatively regulating the Wnt pathway].Med Sci (Paris). 2007 Mar;23(3):243-4. doi: 10.1051/medsci/2007233243. Med Sci (Paris). 2007. PMID: 17349278 French. No abstract available.
-
[Molecular mechanisms of endoderm formation in vertebrates].Tanpakushitsu Kakusan Koso. 2007 Feb;52(2):105-11. Tanpakushitsu Kakusan Koso. 2007. PMID: 17297862 Review. Japanese. No abstract available.
-
[Activation of the beta-catenin-independent pathway by Wnt and the diseases due to its abnormality].Tanpakushitsu Kakusan Koso. 2009 Jan;54(1):1-10. Tanpakushitsu Kakusan Koso. 2009. PMID: 19195220 Review. Japanese. No abstract available.
Cited by
-
Analysis of the dynamic co-expression network of heart regeneration in the zebrafish.Sci Rep. 2016 May 31;6:26822. doi: 10.1038/srep26822. Sci Rep. 2016. PMID: 27241320 Free PMC article.
-
Adipose-Derived Stem Cells Spontaneously Express Neural Markers When Grown in a PEG-Based 3D Matrix.Int J Mol Sci. 2023 Jul 28;24(15):12139. doi: 10.3390/ijms241512139. Int J Mol Sci. 2023. PMID: 37569515 Free PMC article.
-
Multivariate gene expression analysis reveals functional connectivity changes between normal/tumoral prostates.BMC Syst Biol. 2008 Dec 5;2:106. doi: 10.1186/1752-0509-2-106. BMC Syst Biol. 2008. PMID: 19055846 Free PMC article.
-
METTL3/YTDHF1 Stabilizes CSRP1 mRNA to Regulate Glycolysis and Promote Acute Myeloid Leukemia Progression.Cell Biochem Biophys. 2025 Jun;83(2):1993-2007. doi: 10.1007/s12013-024-01610-4. Epub 2024 Nov 20. Cell Biochem Biophys. 2025. PMID: 39565517
-
Haemodynamically dependent valvulogenesis of zebrafish heart is mediated by flow-dependent expression of miR-21.Nat Commun. 2013;4:1978. doi: 10.1038/ncomms2978. Nat Commun. 2013. PMID: 23748970 Free PMC article.
References
-
- Driever W. Curr Opin Genet Dev. 1995;5:610–618. - PubMed
-
- Myers DC, Sepich DS, Solnica-Krezel L. Trends Genet. 2002;18:447–455. - PubMed
-
- Heisenberg CP, Tada M, Rauch GJ, Saude L, Concha ML, Geisler R, Stemple DL, Smith JC, Wilson SW. Nature. 2000;405:76–81. - PubMed
-
- Kilian B, Mansukoski H, Barbosa FC, Ulrich F, Tada M, Heisenberg CP. Mech Dev. 2003;120:467–476. - PubMed
-
- Rauch GJ, Hammerschmidt M, Blader P, Schauerte HE, Strahle U, Ingham PW, McMahon AP, Haffter P. Cold Spring Harbor Symp Quant Biol. 1997;62:227–234. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
