

Distribution of neurons expressing tyrosine hydroxylase in the human cerebral cortex
- PMID: 17593221
- PMCID: PMC2375770
- DOI: 10.1111/j.1469-7580.2007.00760.x
Distribution of neurons expressing tyrosine hydroxylase in the human cerebral cortex
Abstract
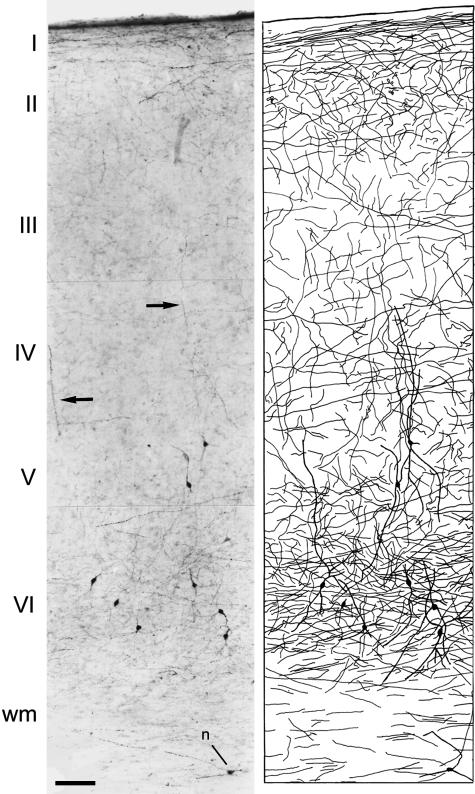
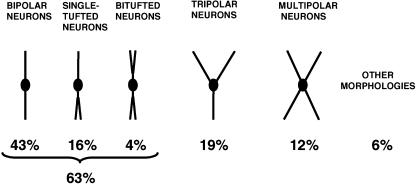
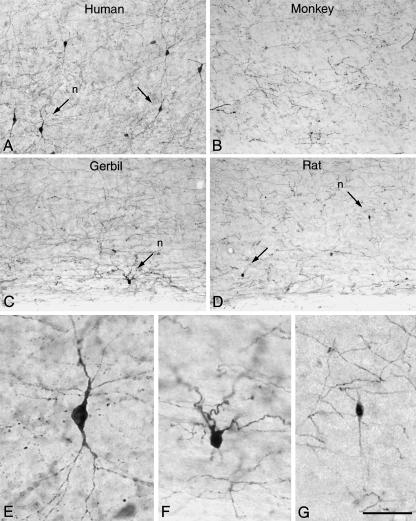
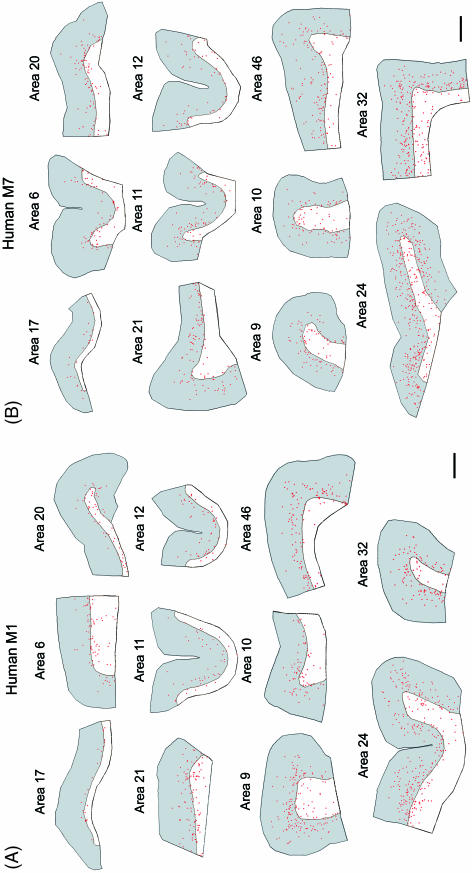
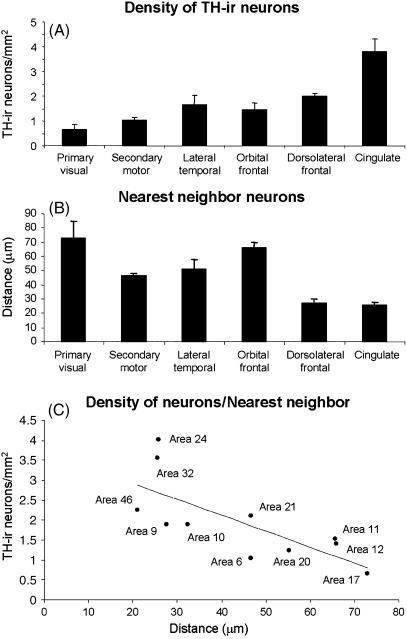
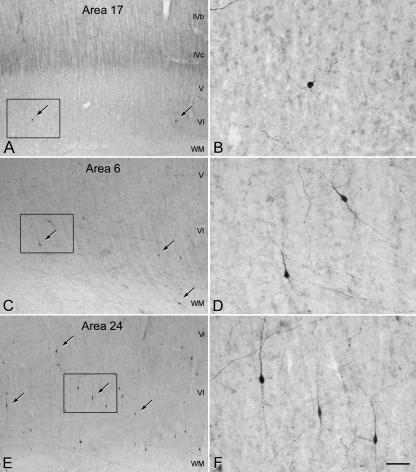
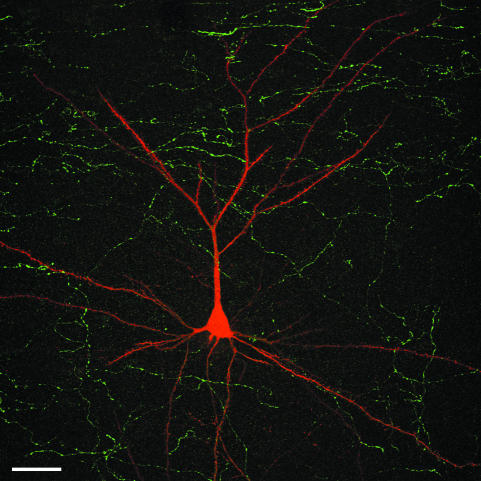
Since the very first detailed description of the different types of cortical interneurons by Cajal, the tremendous variation in the morphology, physiology and neurochemical properties of these cells has become apparent. However, it still remains unclear whether all types of interneurons are present in all cortical areas and species. Here we have focused on tyrosine hydroxylase (TH)-immunoreactive cortical interneurons, which although only present in certain species, are particularly abundant in the human neocortex. We argue that this type of interneuron is more widespread in the human neocortex than in any other species examined so far and that, therefore, it is probably involved in a larger variety of cortical circuits. In addition, notable regional variation can be seen in relation to these interneurons. These differences further emphasize the variability in the design of microcircuits between cortical areas and species, and they probably reflect an evolutionary adaptation of cortical circuits to particular functions.
Figures
References
-
- Ballesteros-Yáñez I, Muñoz A, Contreras J, Gonzalez J, Rodriguez-Veiga E, DeFelipe J. The double bouquet cell in the human cerebral cortex and a comparison with other mammals. J Comp Neurol. 2005;486:344–360. - PubMed
-
- Benavides-Piccione R, DeFelipe J. Different populations of tyrosine-hydroxylase-immunoreactive neurons defined by differential expression of nitric oxide synthase in the human temporal cortex. Cereb Cortex. 2003;13:297–307. - PubMed
-
- Benavides-Piccione R, Arellano J, DeFelipe J. Catecholamine innervation of pyramidal neurons in the human temporal cortex. Cereb Cortex. 2005;15:1584–1591. - PubMed
-
- Benes FM, Taylor JB, Cunningham MC. Convergence and plasticity of monoaminergic systems in the medial prefrontal cortex during the postnatal period: implications for the development of psychopathology. Cereb Cortex. 2000;10:1014–1027. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
