

Interaction of thrombin with PAR1 and PAR4 at the thrombin cleavage site
- PMID: 17595115
- PMCID: PMC2553362
- DOI: 10.1021/bi700597p
Interaction of thrombin with PAR1 and PAR4 at the thrombin cleavage site
Abstract
Investigations determined the critical amino acids for alpha-thrombin's interaction with protease-activated receptors 1 and 4 (PAR1 and PAR4, respectively) at the thrombin cleavage site. Recombinant PAR1 wild-type (wt) exodomain was cleaved by alpha-thrombin with a Km of 28 microM, a kcat of 340 s-1, and a kcat/Km of 1.2 x 10(7). When the P4 or P2 position was mutated to alanine, PAR1-L38A or PAR1-P40A, respectively, the Km was unchanged, 29 or 23 microM, respectively; however, the kcat and kcat/Km were reduced in each case. In contrast, when Asp39 at P3 was mutated to alanine, PAR1-D39A, Km and kcat were both reduced approximately 3-fold, making the kcat/Km the same as that of PAR1-wt exodomain. Recombinant PAR4-wt exodomain was cleaved by alpha-thrombin with a Km of 61 microM, a kcat of 17 s-1, and a kcat/Km of 2.8 x 10(5). When the P5 or P4 position was mutated to alanine, PAR4-L43A or PAR4-P44A, respectively, there was no change in the Km (69 or 56 microM, respectively); however, the kcat was lowered in each case (9.7 or 7.7 s-1, respectively). Mutation of the P2 position (PAR4-P46A) also had no effect on the Km but markedly lowered the kcat and kcat/Km approximately 35-fold. PAR1-wt exodomain and P4 and P3 mutants were noncompetitive inhibitors of alpha-thrombin hydrolyzing Sar-Pro-Arg-pNA. However, PAR1-P40A displayed a mixed type of inhibition. Mutation of P4, P3, or P2 had no effect on the Ki. All PAR4 exodomains were competitive inhibitors of alpha-thrombin. Mutation of P5, P4, or P2 had no effect on the Ki. These investigations show that Leu at P4 in PAR1 or P5 in PAR4 critically influences the kinetics of alpha-thrombin binding and cleavage of PAR1 and PAR4 exodomains. It also implies that factors other than the hirudin-like binding region on PAR1 exodomain predominate in influencing PAR1 cleavage on cells.
Figures
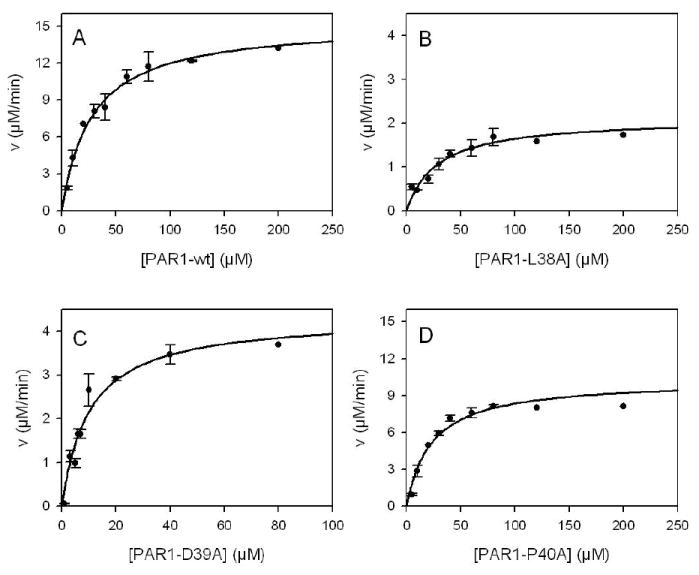


 ) μM PAR1-wt exodomain (A), PAR1-L38A (B), PAR1-D39A (C) or PAR1-P40A (D). Initial velocities data were fit to 8 models of enzyme inhibition using a global analysis nonlinear least squares regression analysis to determine the Ki and the type of inhibition (see Methods). Double reciprocal plots (insets) are shown only for graphical representation of the model and were not used to determine the Ki or type of inhibition.
) μM PAR1-wt exodomain (A), PAR1-L38A (B), PAR1-D39A (C) or PAR1-P40A (D). Initial velocities data were fit to 8 models of enzyme inhibition using a global analysis nonlinear least squares regression analysis to determine the Ki and the type of inhibition (see Methods). Double reciprocal plots (insets) are shown only for graphical representation of the model and were not used to determine the Ki or type of inhibition.
 ), 100 (▼), 120 (∇) or 200 (○) μM PAR4-wt (A), PAR4-L43A (B), PAR4-P44A (C), or PAR4-P46A (D). Initial velocities data were fit to 8 models of enzyme inhibition using a global analysis nonlinear least squares regression analysis to determine the Ki and the type of inhibition (see Methods). Double reciprocal plots (insets) are shown only for graphical representation of the model and were not used to determine the Ki or type of inhibition.
), 100 (▼), 120 (∇) or 200 (○) μM PAR4-wt (A), PAR4-L43A (B), PAR4-P44A (C), or PAR4-P46A (D). Initial velocities data were fit to 8 models of enzyme inhibition using a global analysis nonlinear least squares regression analysis to determine the Ki and the type of inhibition (see Methods). Double reciprocal plots (insets) are shown only for graphical representation of the model and were not used to determine the Ki or type of inhibition.References
-
- Mann KG, Brummel K, Butenas S. What is all that thrombin for? J Thromb Haemost. 2003;1:1504–1514. - PubMed
-
- Coughlin SR. Thrombin signalling and protease-activated receptors. Nature. 2000;407:258–264. - PubMed
-
- Swift S, Sheridan PJ, Covic L, Kuliopulos A. PAR1 thrombin receptor-G protein interactions. Separation of binding and coupling determinants in the galpha subunit. J Biol Chem. 2000;275:2627–2635. - PubMed
-
- Verrall S, Ishii M, Chen M, Wang L, Tram T, Coughlin SR. The thrombin receptor second cytoplasmic loop confers coupling to Gq-like G proteins in chimeric receptors. Additional evidence for a common transmembrane signaling and G protein coupling mechanism in G protein-coupled receptors. J Biol Chem. 1997;272:6898–6902. - PubMed
-
- Steinhoff M, Buddenkotte J, Shpacovitch V, Rattenholl A, Moormann C, Vergnolle N, Luger TA, Hollenberg MD. Proteinase-activated receptors: transducers of proteinase-mediated signaling in inflammation and immune response. Endocr Rev. 2005;26:1–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
