

Presence of thiamine pyrophosphate in mammalian peroxisomes
- PMID: 17596263
- PMCID: PMC1913525
- DOI: 10.1186/1471-2091-8-10
Presence of thiamine pyrophosphate in mammalian peroxisomes
Abstract
Background: Thiamine pyrophosphate (TPP) is a cofactor for 2-hydroxyacyl-CoA lyase 1 (HACL1), a peroxisomal enzyme essential for the alpha-oxidation of phytanic acid and 2-hydroxy straight chain fatty acids. So far, HACL1 is the only known peroxisomal TPP-dependent enzyme in mammals. Little is known about the transport of metabolites and cofactors across the peroxisomal membrane and no peroxisomal thiamine or TPP carrier has been identified in mammals yet. This study was undertaken to get a better insight into these issues and to shed light on the role of TPP in peroxisomal metabolism.
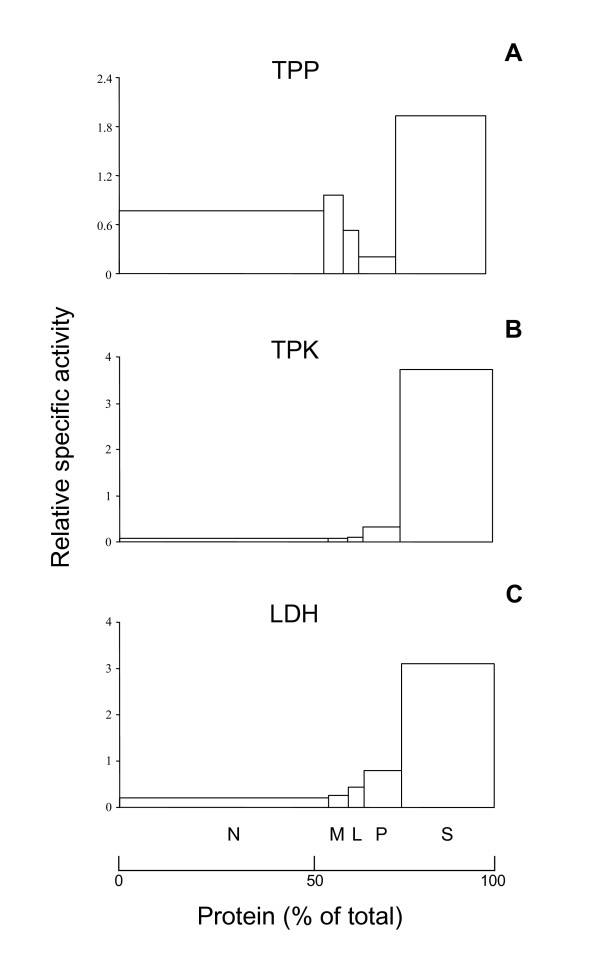
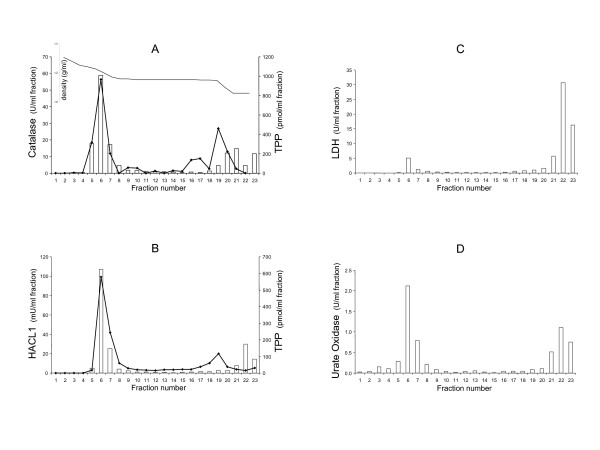
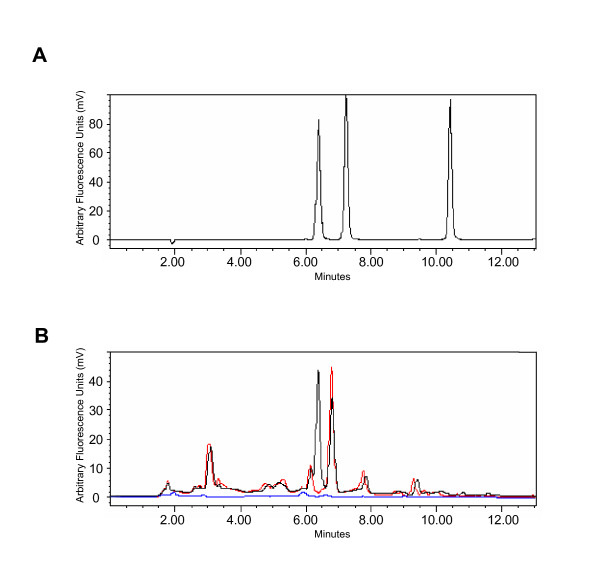
Results: Because of the crucial role of the cofactor TPP, we reanalyzed its subcellular localization in rat liver. In addition to the known mitochondrial and cytosolic pools, we demonstrated, for the first time, that peroxisomes contain TPP (177 +/- 2 pmol/mg protein). Subsequently, we verified whether TPP could be synthesized from its precursor thiamine, in situ, by a peroxisomal thiamine pyrophosphokinase (TPK). However, TPK activity was exclusively recovered in the cytosol.
Conclusion: Our results clearly indicate that mammalian peroxisomes do contain TPP but that no pyrophosphorylation of thiamine occurs in these organelles, implying that thiamine must enter the peroxisome already pyrophosphorylated. Consequently, TPP entry may depend on a specific transport system or, in a bound form, on HACL1 translocation.
Figures
Similar articles
-
The role of 2-hydroxyacyl-CoA lyase, a thiamin pyrophosphate-dependent enzyme, in the peroxisomal metabolism of 3-methyl-branched fatty acids and 2-hydroxy straight-chain fatty acids.Biochem Soc Trans. 2007 Nov;35(Pt 5):876-80. doi: 10.1042/BST0350876. Biochem Soc Trans. 2007. PMID: 17956236 Review.
-
Role of thiamine pyrophosphate in oligomerisation, functioning and import of peroxisomal 2-hydroxyacyl-CoA lyase.Biochim Biophys Acta. 2011 Oct;1814(10):1226-33. doi: 10.1016/j.bbapap.2011.06.007. Epub 2011 Jun 25. Biochim Biophys Acta. 2011. PMID: 21708296
-
[Free and bound thiamine pyrophosphate level in rat liver mitochondria in various saturation of the body with thiamine].Vopr Med Khim. 1981 Mar-Apr;27(2):239-43. Vopr Med Khim. 1981. PMID: 7281564 Russian.
-
Thiamine pyrophosphate biosynthesis and transport in the nematode Caenorhabditis elegans.Genetics. 2004 Oct;168(2):845-54. doi: 10.1534/genetics.104.028605. Genetics. 2004. PMID: 15514058 Free PMC article.
-
Protein translocation machineries: how organelles bring in matrix proteins.FEMS Yeast Res. 2005 Nov;5(11):1037-45. doi: 10.1016/j.femsyr.2005.03.004. FEMS Yeast Res. 2005. PMID: 16269392 Review.
Cited by
-
The effect of thiamine and thiamine pyrophosphate on oxidative liver damage induced in rats with cisplatin.Biomed Res Int. 2013;2013:783809. doi: 10.1155/2013/783809. Epub 2013 Jun 6. Biomed Res Int. 2013. PMID: 23841092 Free PMC article.
-
Metabolic and structural role of thiamine in nervous tissues.Cell Mol Neurobiol. 2008 Nov;28(7):923-31. doi: 10.1007/s10571-008-9297-7. Epub 2008 Jul 19. Cell Mol Neurobiol. 2008. PMID: 18642074 Free PMC article. Review.
-
Transketolase (TKT) activity and nuclear localization promote hepatocellular carcinoma in a metabolic and a non-metabolic manner.J Exp Clin Cancer Res. 2019 Apr 11;38(1):154. doi: 10.1186/s13046-019-1131-1. J Exp Clin Cancer Res. 2019. PMID: 30971297 Free PMC article.
-
The Thiamin-Requiring 3 Mutation of Arabidopsis 5-Deoxyxylulose-Phosphate Synthase 1 Highlights How the Thiamin Economy Impacts the Methylerythritol 4-Phosphate Pathway.Front Plant Sci. 2021 Aug 6;12:721391. doi: 10.3389/fpls.2021.721391. eCollection 2021. Front Plant Sci. 2021. PMID: 34421975 Free PMC article.
-
Thiamine deficiency in pregnancy and lactation: implications and present perspectives.Front Nutr. 2023 Apr 20;10:1080611. doi: 10.3389/fnut.2023.1080611. eCollection 2023. Front Nutr. 2023. PMID: 37153911 Free PMC article. Review.
References
-
- Foulon V, Antonenkov VD, Croes K, Waelkens E, Mannaerts GP, Van Veldhoven PP, Casteels M. Purification, molecular cloning, and expression of 2-hydroxyphytanoyl-CoA lyase, a peroxisomal thiamine pyrophosphate-dependent enzyme that catalyzes the carbon-carbon bond cleavage during alpha-oxidation of 3-methyl-branched fatty acids. Proc Natl Acad Sci U S A. 1999;96:10039–10044. doi: 10.1073/pnas.96.18.10039. - DOI - PMC - PubMed
-
- Sniekers M, Foulon V, Mannaerts GP, Van Maldergem L, Mandel H, Gelb BD, Casteels M, Van Veldhoven PP. Thiamine pyrophosphate: an essential cofactor for the alpha-oxidation in mammals--implications for thiamine deficiencies? Cell Mol Life Sci. 2006;63:1553–1563. doi: 10.1007/s00018-005-5603-4. - DOI - PMC - PubMed
-
- Foulon V, Sniekers M, Huysmans E, Asselberghs S, Mahieu V, Mannaerts GP, Van Veldhoven PP, Casteels M. Breakdown of 2-hydroxylated straight chain fatty acids via peroxisomal 2-hydroxyphytanoyl-CoA lyase: a revised pathway for the alpha-oxidation of straight chain fatty acids. J Biol Chem. 2005;280:9802–9812. doi: 10.1074/jbc.M413362200. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
