

Role of auditory cortex in sound localization in the midsagittal plane
- PMID: 17596417
- PMCID: PMC7116534
- DOI: 10.1152/jn.00444.2007
Role of auditory cortex in sound localization in the midsagittal plane
Abstract
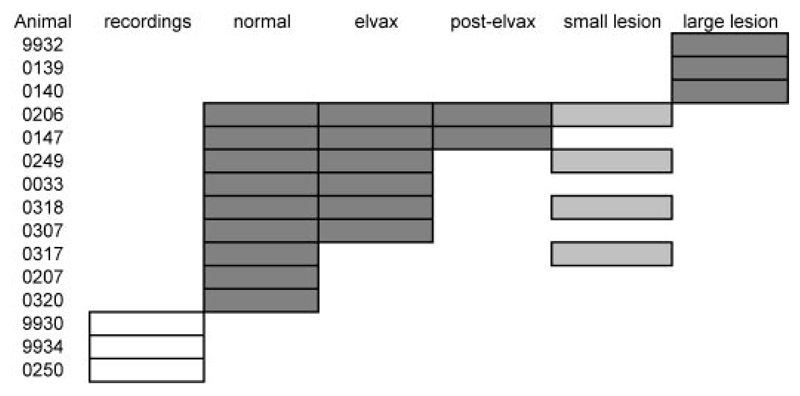
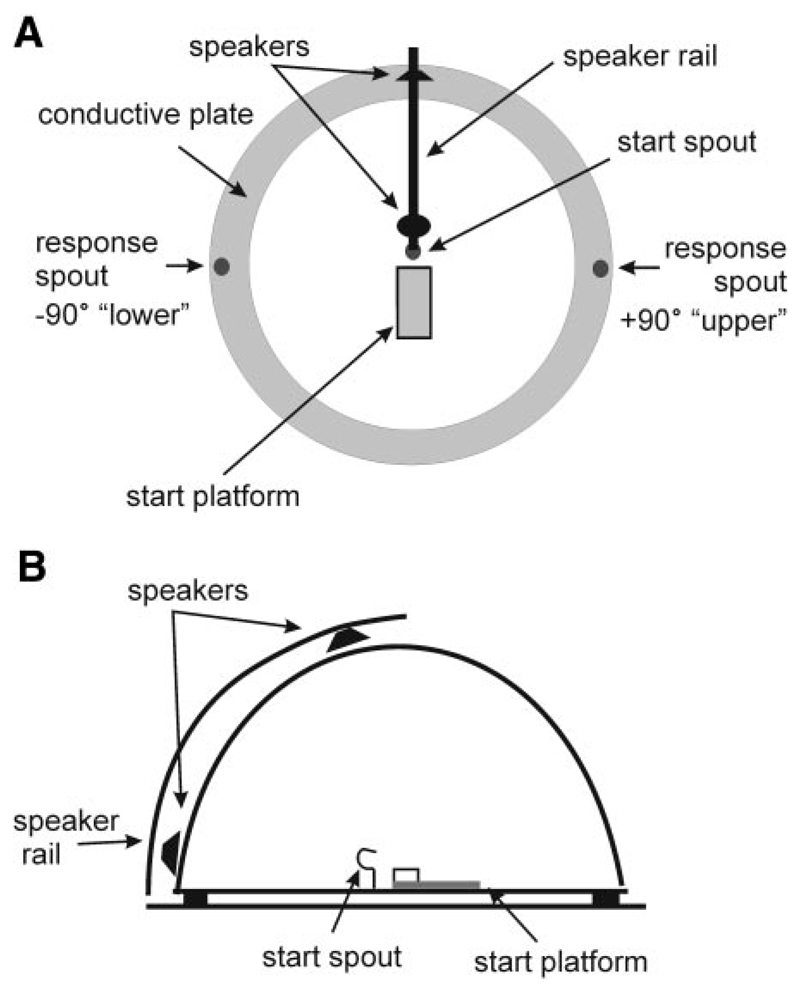
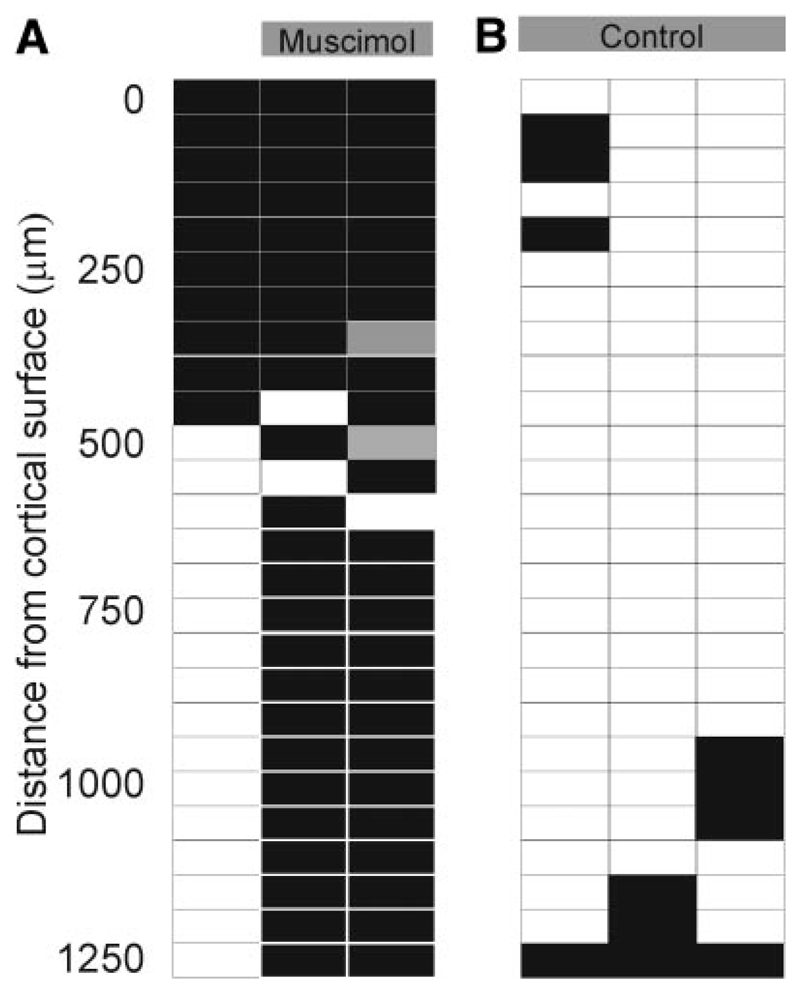
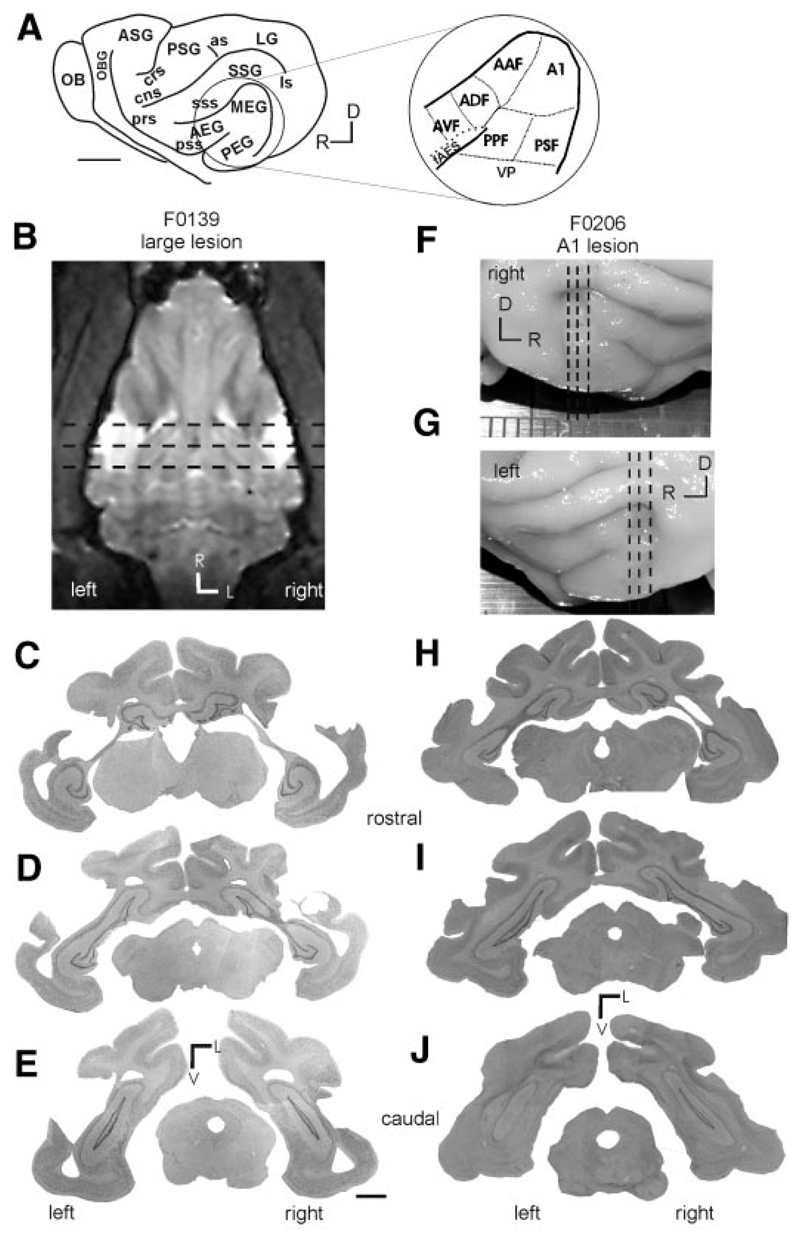
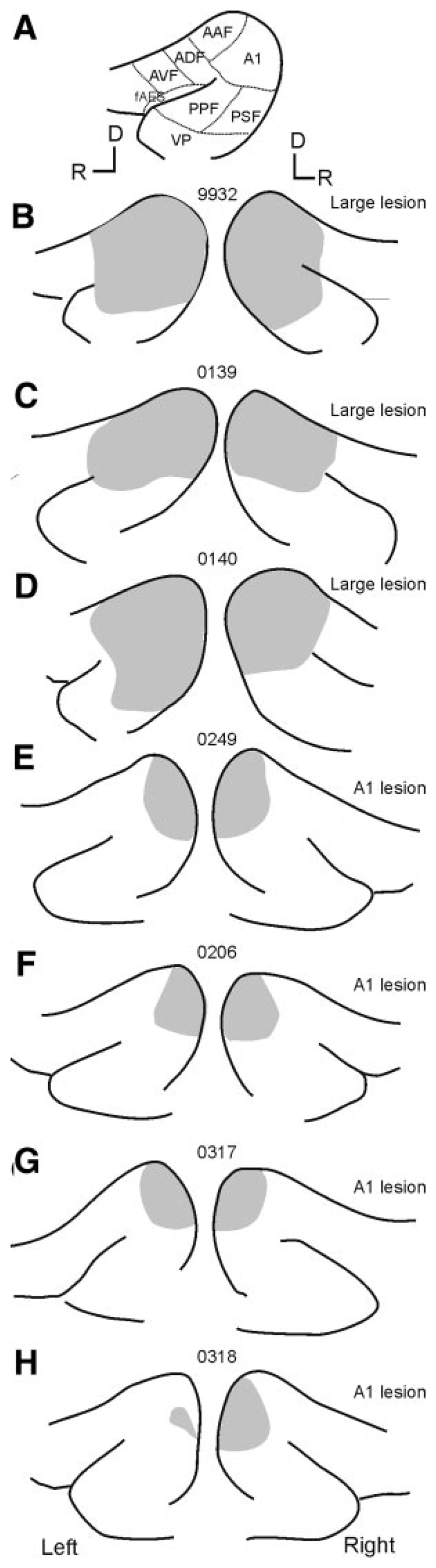
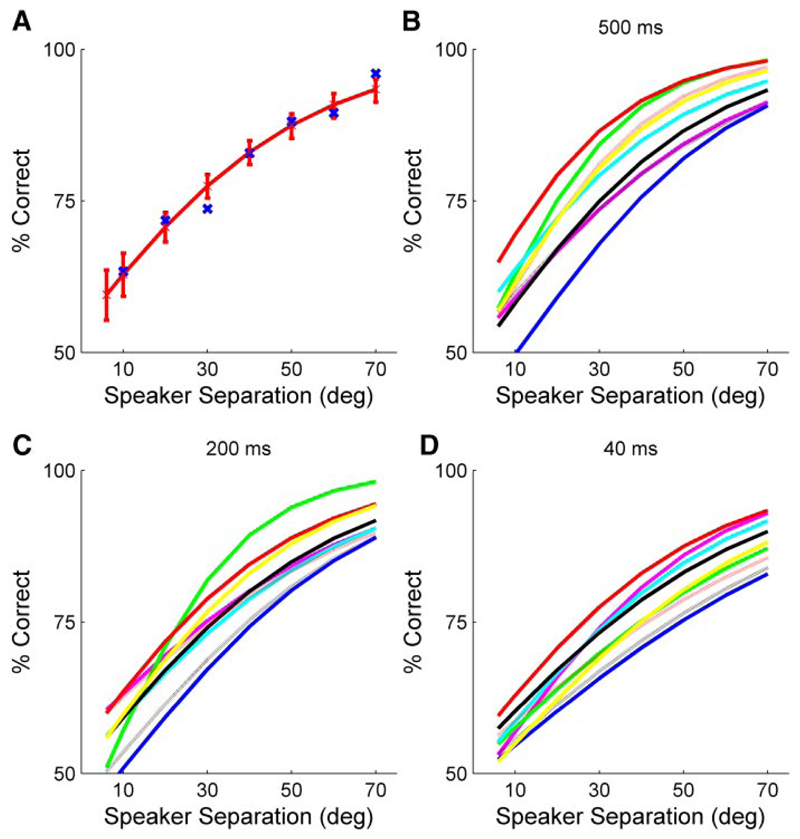
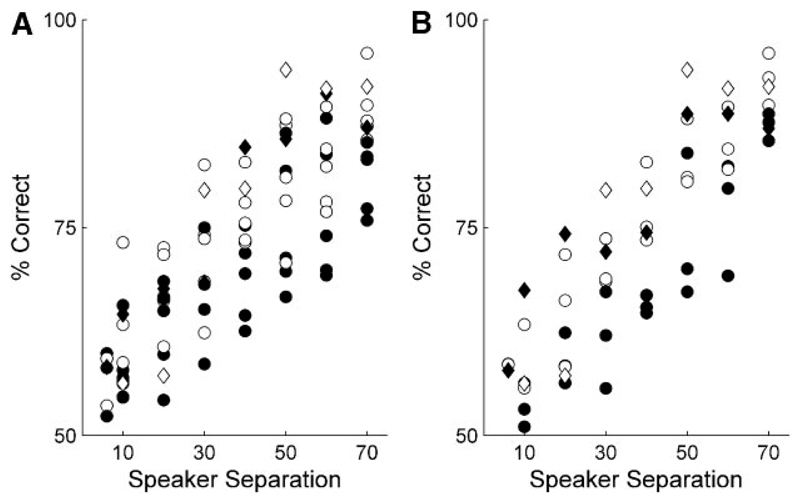
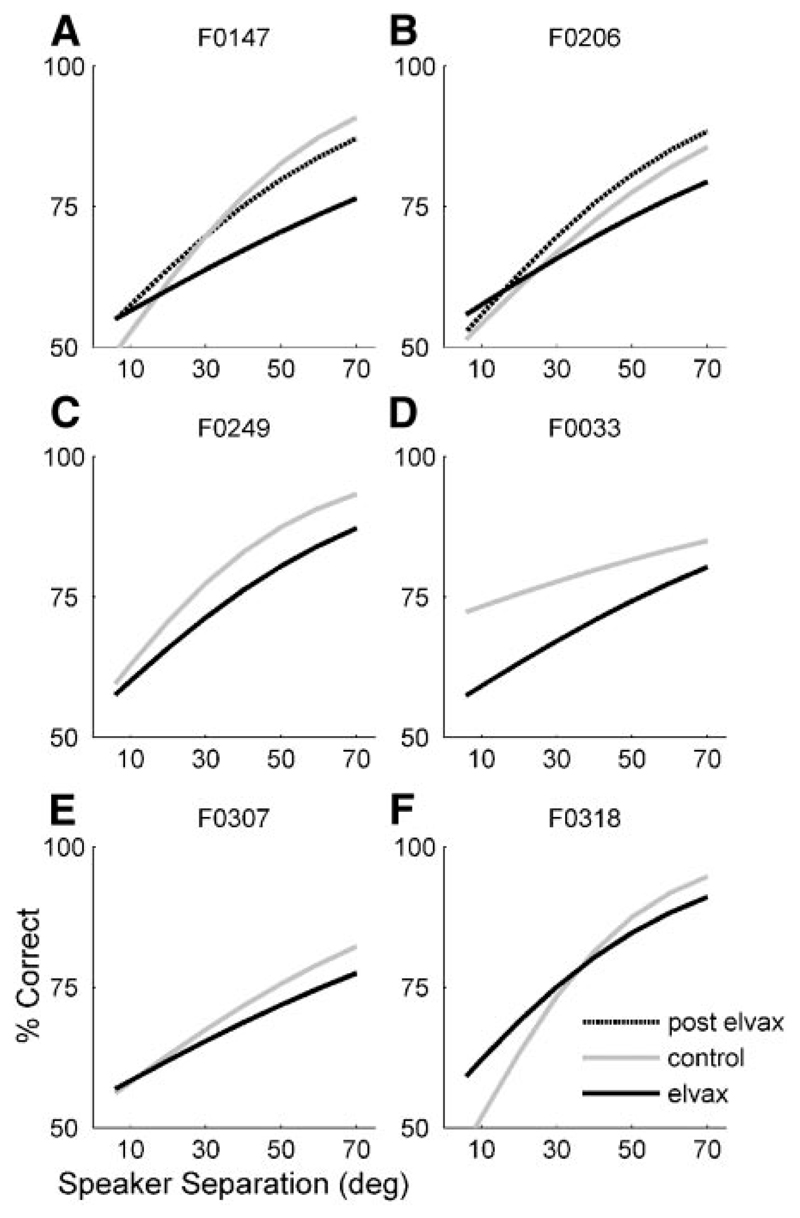
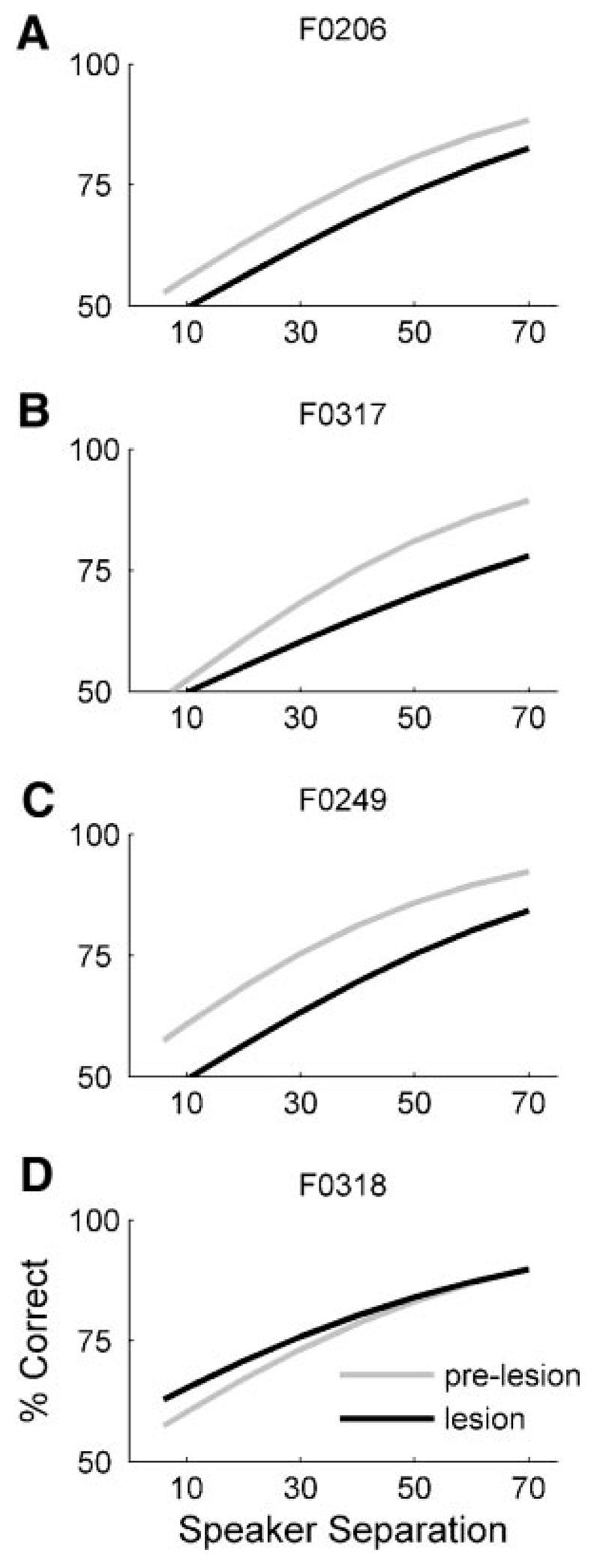
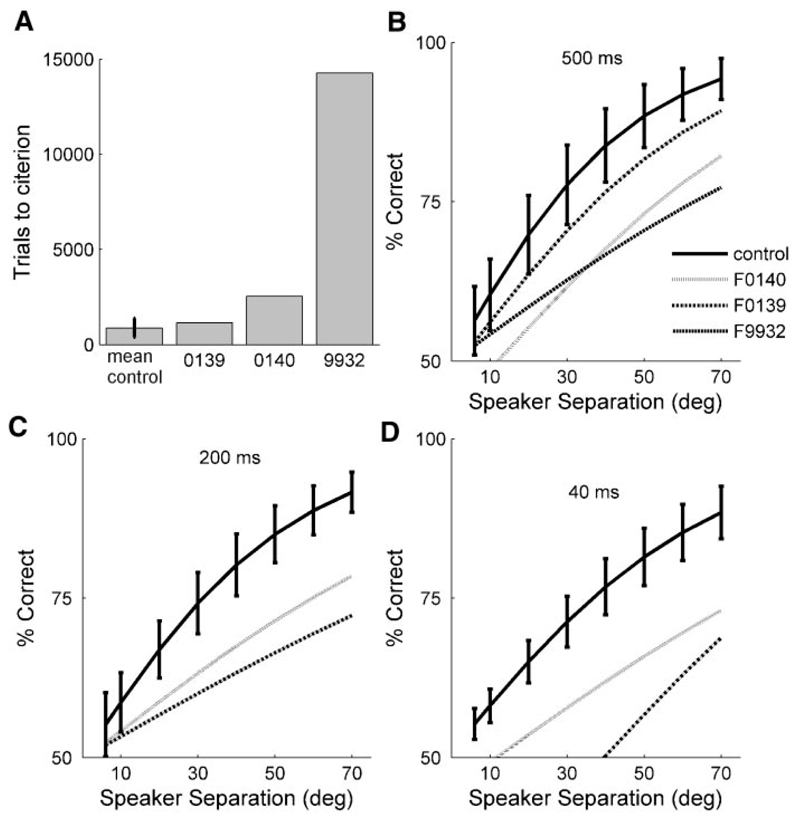
Although the auditory cortex is known to be essential for normal sound localization in the horizontal plane, its contribution to vertical localization has not so far been examined. In this study, we measured the acuity with which ferrets could discriminate between two speakers in the midsagittal plane before and after silencing activity bilaterally in the primary auditory cortex (A1). This was achieved either by subdural placement of Elvax implants containing the GABA A receptor agonist muscimol or by making aspiration lesions after determining the approximate location of A1 electrophysiologically. Psychometric functions and minimum audible angles were measured in the upper hemifield for 500-, 200-, and 40-ms noise bursts. Muscimol-Elvax inactivation of A1 produced a small but significant deficit in the animals' ability to localize brief (40-ms) sounds, which was reversed after removal of the Elvax implants. A similar deficit in vertical localization was observed after bilateral aspiration lesions of A1, whereas performance at longer sound durations was unaffected. Another group of ferrets received larger lesions, encompassing both primary and nonprimary auditory cortical areas, and showed a greater deficit with performance being impaired for long- and short-duration (500- and 40-ms, respectively) stimuli. These data suggest that the integrity of the auditory cortex is required to successfully utilize spectral localization cues, which are thought to provide the basis for vertical localization, and that multiple cortical fields, including A1, contribute to this task.
Figures
References
-
- Beitel RE, Kaas JH. Effects of bilateral and unilateral ablation of auditory cortex in cats on the unconditioned head orienting response to acoustic stimuli. J Neurophysiol. 1993;70:351–369. - PubMed
-
- Berry G, Matthews JNS, Armitage P. Statistical Methods in Medical Research. Oxford, UK: Blackwell Science; 2001.
-
- Bizley JK. Organization and Function of Ferret Auditory Cortex (PhD thesis) Oxford, UK: Univ. of Oxford; 2005.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources