

Quantal and nonquantal transmission in calyx-bearing fibers of the turtle posterior crista
- PMID: 17596419
- PMCID: PMC3397384
- DOI: 10.1152/jn.00332.2007
Quantal and nonquantal transmission in calyx-bearing fibers of the turtle posterior crista
Abstract
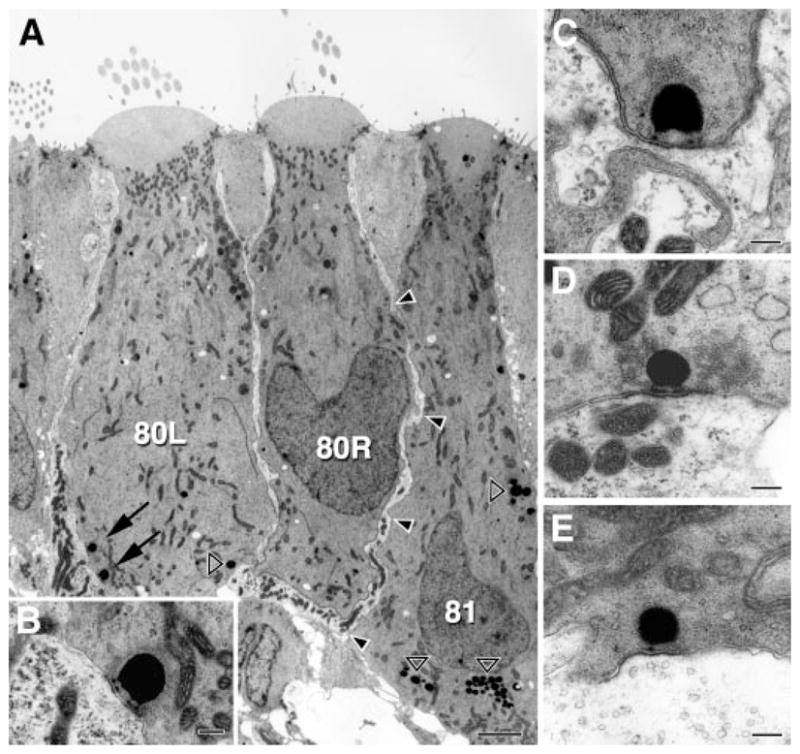
Intracellular recordings were made from nerve fibers in the posterior ampullary nerve near the neuroepithelium. Calyx-bearing afferents were identified by their distinctive efferent-mediated responses. Such fibers receive inputs from both type I and type II hair cells. Type II inputs are made by synapses on the outer face of the calyx ending and on the boutons of dimorphic fibers. Quantal activity, consisting of brief mEPSPs, is reduced by lowering the external concentration of Ca2+ and blocked by the AMPA-receptor antagonist CNQX. Poisson statistics govern the timing of mEPSPs, which occur at high rates (250-2,500/s) in the absence of mechanical stimulation. Excitation produced by canal-duct indentation can increase mEPSP rates to nearly 5,000/s. As the rate increases, mEPSPs can change from a monophasic depolarization to a biphasic depolarizing-hyperpolarizing sequence, both of whose components are blocked by CNQX. Blockers of voltage-gated currents affect mEPSP size, which is decreased by TTX and is increased by linopirdine. mEPSP size decreases severalfold after impalement. The size decrease, although it may be triggered by the depolarization occurring during impalement, persists even at hyperpolarized membrane potentials. Nonquantal transmission is indicated by shot-noise calculations and by the presence of voltage modulations after quantal activity is abolished pharmacologically. An ultrastructural study shows that inner-face inputs from type I hair cells outnumber outer-face inputs from type II hair cells by an almost 6:1 ratio.
Figures












References
-
- Baird IL. Anatomical features of the inner ear in submammalian vertebrates. In: Keidel WD, Neff WD, editors. Handbook of Sensory Physiology. Berlin: Springer-Verlag; 1974. pp. 159–212.
-
- Baird RA, Desmadryl G, Fernández C, Goldberg JM. The vestibular nerve of the chinchilla. II. Relation between afferent response properties and peripheral innervation patterns in the semicircular canals. J Neurophysiol. 1988;60:182–203. - PubMed
-
- Bao H, Wong WH, Goldberg JM, Eatock RA. Voltage-gated calcium channel currents in type I and type II hair cells isolated from the rat crista. J Neurophysiol. 2003;90:155–164. - PubMed
-
- Behrend O, Schwark C, Kunihiro T, Strupp M. Cyclic GMP inhibits and shifts the activation curve of the delayed-rectifier (IK1) of type I mammalian vestibular hair cells. Neuroreport. 1997;8:2687–2690. - PubMed
-
- Boettger T, Hubner CA, Maier H, Rust MB, Beck FX, Jentsch TJ. Deafness and renal tubular acidosis in mice lacking the K-Cl co-transporter KCC4. Nature. 2002;416:874–878. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous