

Serotonin activates the hypothalamic-pituitary-adrenal axis via serotonin 2C receptor stimulation
- PMID: 17596444
- PMCID: PMC6672238
- DOI: 10.1523/JNEUROSCI.2584-06.2007
Serotonin activates the hypothalamic-pituitary-adrenal axis via serotonin 2C receptor stimulation
Abstract
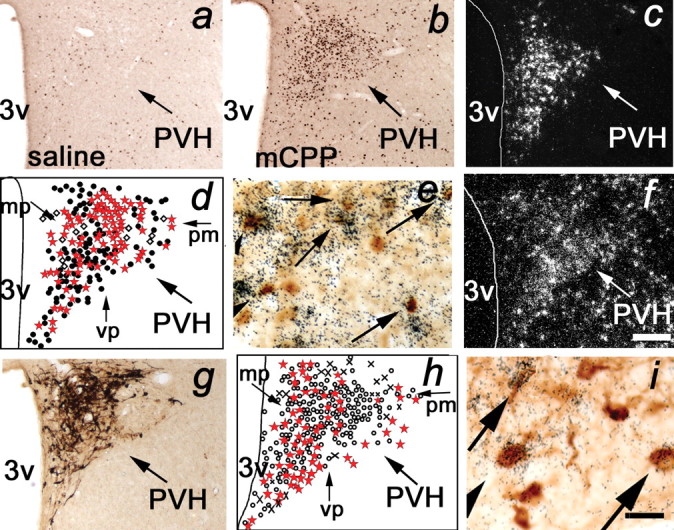
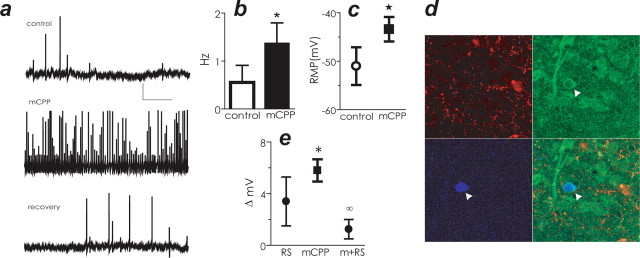
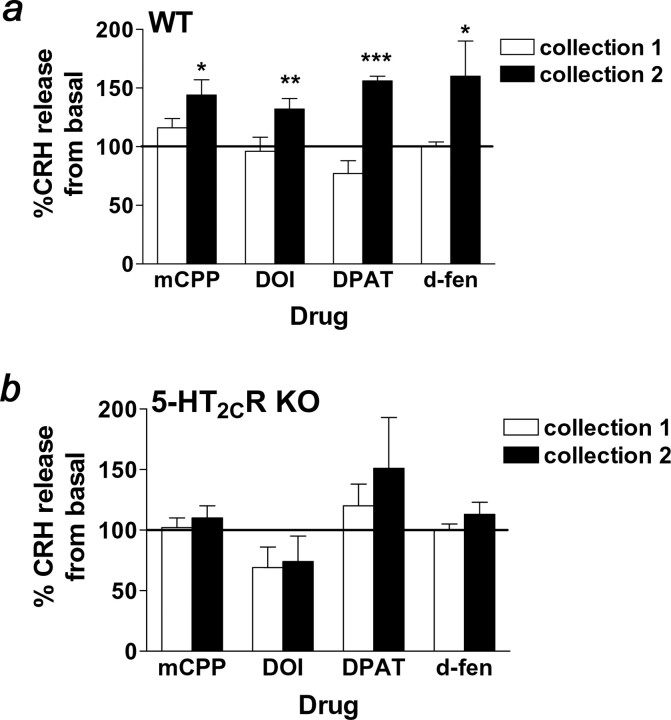
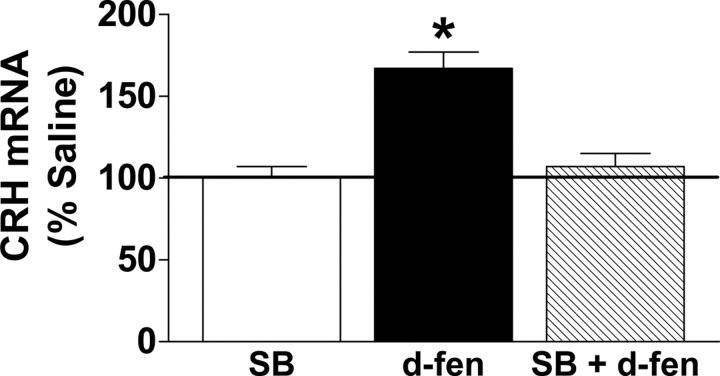
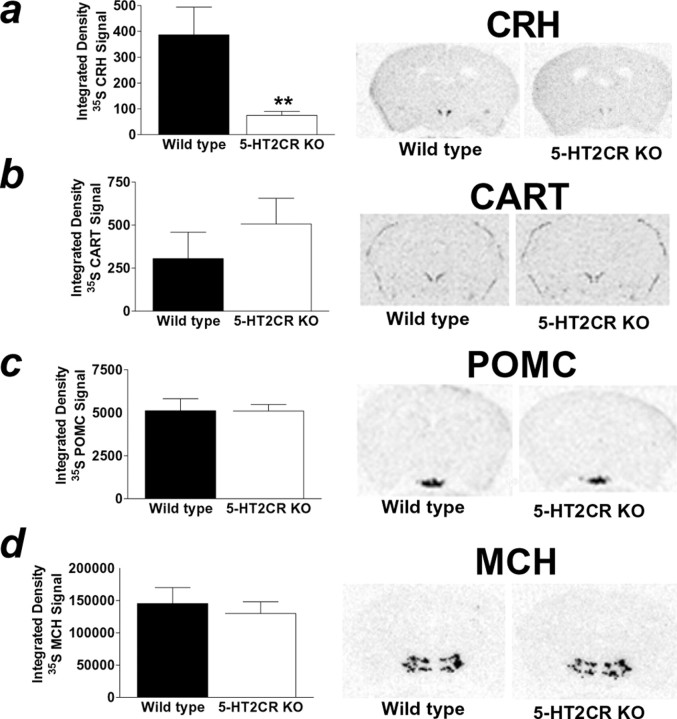
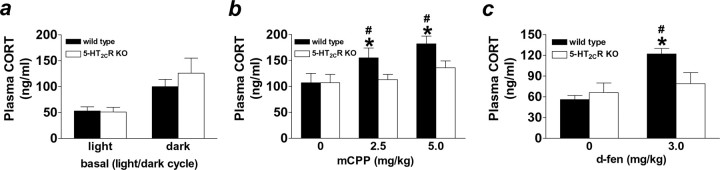
The dynamic interplay between serotonin [5-hydroxytryptamine (5-HT)] neurotransmission and the hypothalamic-pituitary-adrenal (HPA) axis has been extensively studied over the past 30 years, but the underlying mechanism of this interaction has not been defined. A possibility receiving little attention is that 5-HT regulates upstream corticotropin-releasing hormone (CRH) signaling systems via activation of serotonin 2C receptors (5-HT(2C)Rs) in the paraventricular nucleus of the hypothalamus (PVH). Through complementary approaches in wild-type rodents and 5-HT(2C)R-deficient mice, we determined that 5-HT(2C)Rs are necessary for 5-HT-induced HPA axis activation. We used laser-capture PVH microdissection followed by microarray analysis to compare the expression of 13 5-HTRs. Only 5-HT(2C)R and 5-HT(1D)R transcripts were consistently identified as present in the PVH, and of these, the 5-HT(2C)R was expressed at a substantially higher level. The abundant expression of 5-HT(2C)Rs in the PVH was confirmed with in situ hybridization histochemistry. Dual-neurohistochemical labeling revealed that approximately one-half of PVH CRH-containing neurons coexpressed 5-HT(2C)R mRNA. We observed that PVH CRH neurons consistently depolarized in the presence of a high-affinity 5-HT(2C)R agonist, an effect blocked by a 5-HT(2C)R antagonist. Supporting the importance of 5-HT(2C)Rs in CRH neuronal activity, genetic inactivation of 5-HT(2C)Rs produced a downregulation of CRH mRNA and blunted CRH and corticosterone release after 5-HT compound administration. These findings thus provide a mechanistic explanation for the longstanding observation of HPA axis stimulation in response to 5-HT and thereby give insight into the neural circuitry mediating the complex neuroendocrine responses to stress.
Figures
References
-
- Bagdy G, Calogero AE, Aulakh CS, Szemeredi K, Murphy DL. Long-term cortisol treatment impairs behavioral and neuroendocrine responses to 5-HT1 agonists in the rat. Neuroendocrinology. 1989;50:241–247. - PubMed
-
- Bhatnagar S, Sun LM, Raber J, Maren S, Julius D, Dallman MF. Changes in anxiety-related behaviors and hypothalamic-pituitary-adrenal activity in mice lacking the 5-HT-3A receptor. Physiol Behav. 2004;81:545–555. - PubMed
-
- Broocks A, Bandelow B, George A, Jestrabeck C, Opitz M, Bartmann U, Gleiter CH, Meineke I, Roed IS, Ruther E, Hajak G. Increased psychological responses and divergent neuroendocrine responses to m-CPP and ipsapirone in patients with panic disorder. Int Clin Psychopharmacol. 2000;15:153–161. - PubMed
-
- Calogero AE, Bernardini R, Margioris AN, Bagdy G, Gallucci WT, Munson PJ, Tamarkin L, Tomai TP, Brady L, Gold PW, Chrousos GP. Effects of serotonergic agonists and antagonists on corticotropin-releasing hormone secretion by explanted rat hypothalami. Peptides. 1989;10:189–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous