

Mechanisms in the regulation of aromatase in developing ovary and placenta
- PMID: 17596931
- PMCID: PMC2075360
- DOI: 10.1016/j.jsbmb.2007.05.001
Mechanisms in the regulation of aromatase in developing ovary and placenta
Abstract
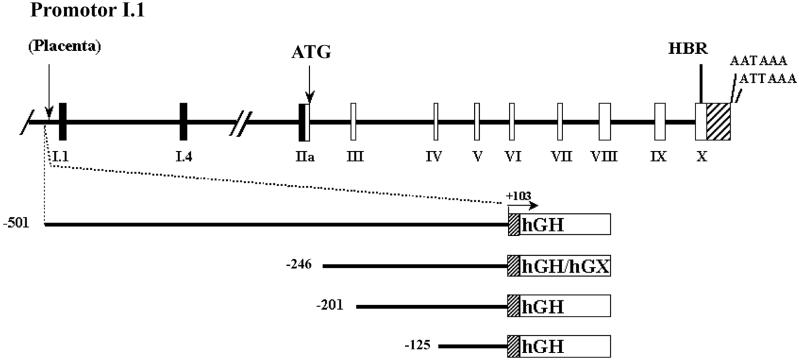
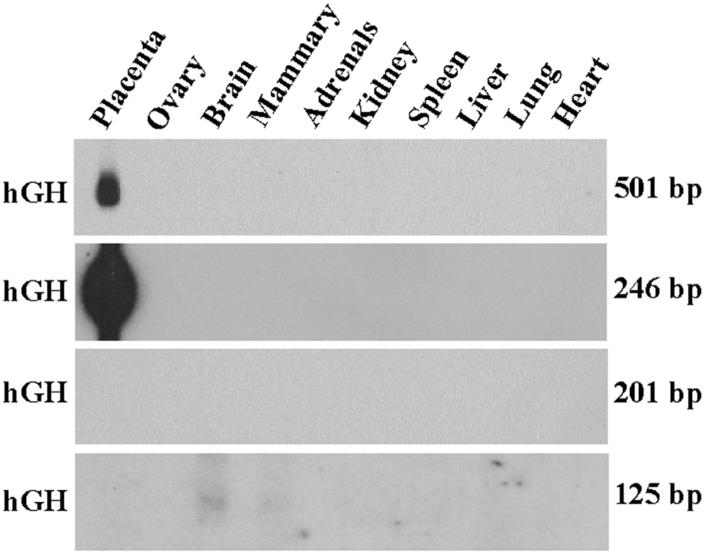
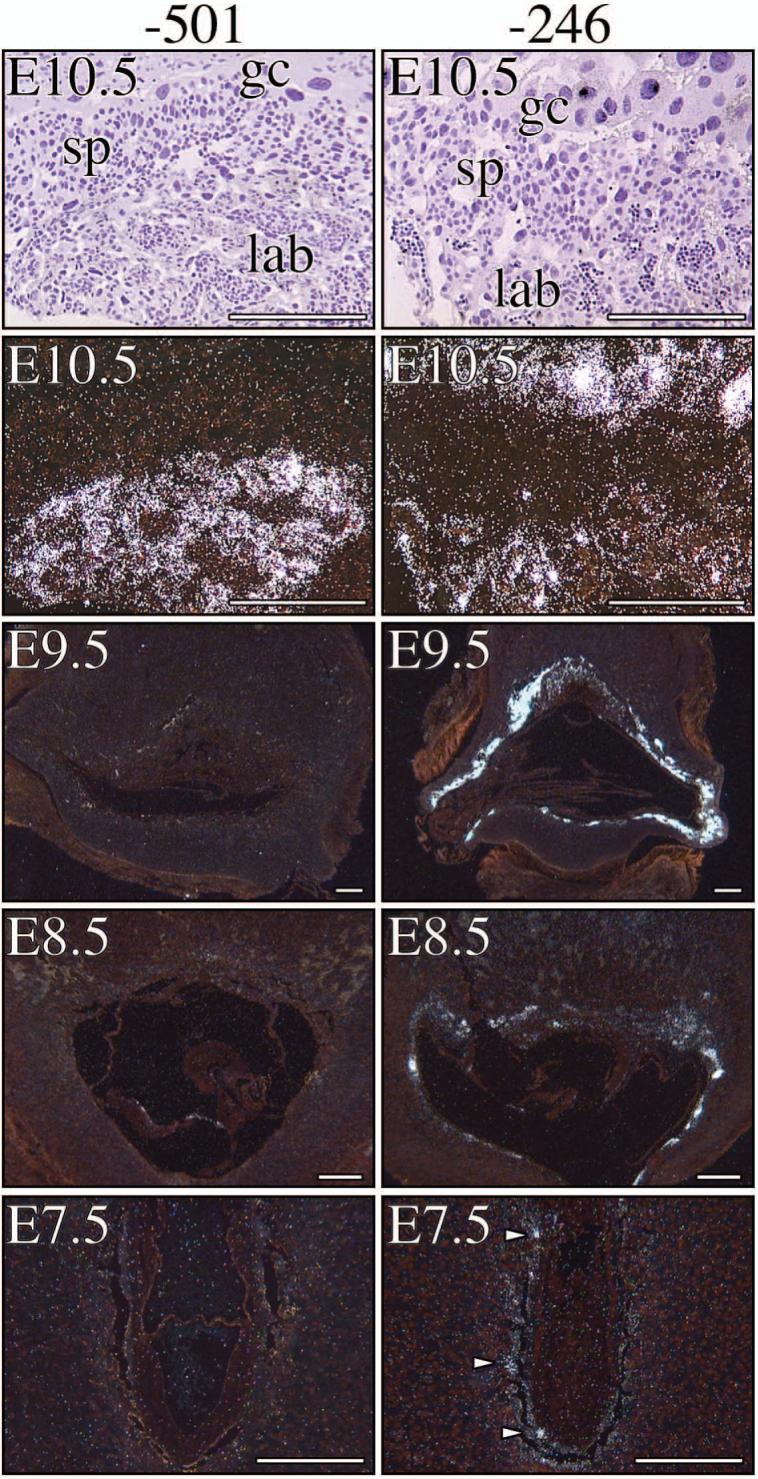
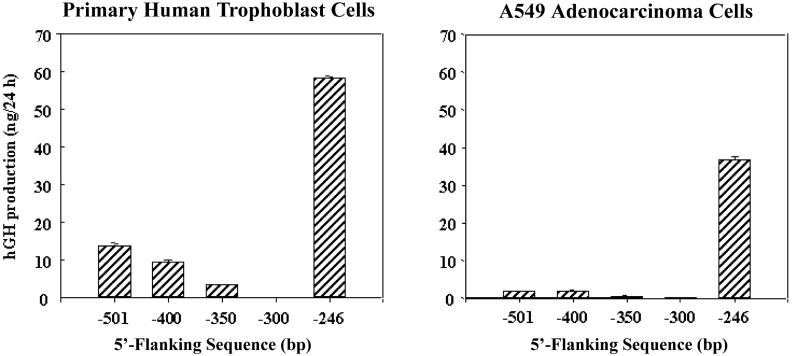
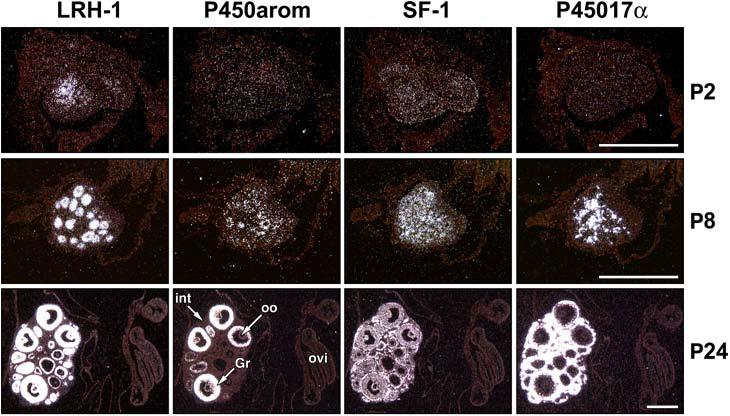
During human gestation, the placental syncytiotrophoblast develops the capacity to synthesize large amounts of estrogen from C(19)-steroids secreted by the fetal adrenals. The conversion of C(19)-steroids to estrogens is catalyzed by aromatase P450 (P450arom), product of the CYP19 gene. The placenta-specific promoter of the hCYP19 gene lies approximately 100,000 bp upstream of the translation initiation site in exon II. In studies using transgenic mice and transfected human trophoblast cells we have defined a 246-bp region upstream of placenta-specific exon I.1 that mediates placental cell-specific expression. Using transgenic mice, we also observed that as little as 278 bp of DNA flanking the 5'-end of ovary-specific hCYP19 exon IIa was sufficient to target ovary-specific expression. This ovary-specific promoter contains response elements that bind cAMP-response element-binding protein (CREB) and the orphan nuclear receptors SF-1 and LRH-1, which are required for cAMP-mediated stimulation of CYP19 expression in granulosa and luteal cells during the estrous cycle and pregnancy. In this article, we review our studies to define genomic regions and response elements that mediate placenta-specific expression of the hCYP19 gene. The temporal and spatial expression of LRH-1 versus SF-1 in the developing gonad during mouse embryogenesis and in the postnatal ovary also will be considered.
Figures
Similar articles
-
A 500-bp region, approximately 40 kb upstream of the human CYP19 (aromatase) gene, mediates placenta-specific expression in transgenic mice.Proc Natl Acad Sci U S A. 1999 Apr 13;96(8):4575-80. doi: 10.1073/pnas.96.8.4575. Proc Natl Acad Sci U S A. 1999. PMID: 10200304 Free PMC article.
-
Characterization of the regulatory regions of the human aromatase (P450arom) gene involved in placenta-specific expression.Mol Endocrinol. 1998 Nov;12(11):1764-77. doi: 10.1210/mend.12.11.0190. Mol Endocrinol. 1998. PMID: 9817601
-
Transcriptional regulation of aromatase in placenta and ovary.J Steroid Biochem Mol Biol. 2005 May;95(1-5):25-33. doi: 10.1016/j.jsbmb.2005.04.016. J Steroid Biochem Mol Biol. 2005. PMID: 15961305 Review.
-
Genomic regions that mediate placental cell-specific and developmental regulation of human Cyp19 (aromatase) gene expression in transgenic mice.Endocrinology. 2005 May;146(5):2481-8. doi: 10.1210/en.2004-1606. Epub 2005 Jan 27. Endocrinology. 2005. PMID: 15677755
-
Tissue-specific promoters regulate aromatase cytochrome P450 expression.J Steroid Biochem Mol Biol. 1993 Mar;44(4-6):321-30. doi: 10.1016/0960-0760(93)90235-o. J Steroid Biochem Mol Biol. 1993. PMID: 8476746 Review.
Cited by
-
The bile acid synthesis pathway is present and functional in the human ovary.PLoS One. 2009 Oct 6;4(10):e7333. doi: 10.1371/journal.pone.0007333. PLoS One. 2009. PMID: 19806215 Free PMC article.
-
Molecular regulation of human placental growth factor (PlGF) gene expression in placental villi and trophoblast cells is mediated via the protein kinase a pathway.Reprod Sci. 2011 Mar;18(3):219-28. doi: 10.1177/1933719110389337. Epub 2010 Dec 6. Reprod Sci. 2011. PMID: 21135203 Free PMC article.
-
Neuroprotective actions of brain aromatase.Front Neuroendocrinol. 2009 Jul;30(2):106-18. doi: 10.1016/j.yfrne.2009.04.016. Epub 2009 May 18. Front Neuroendocrinol. 2009. PMID: 19450619 Free PMC article. Review.
-
Transforming growth factor-beta inhibits aromatase gene transcription in human trophoblast cells via the Smad2 signaling pathway.Reprod Biol Endocrinol. 2009 Dec 9;7:146. doi: 10.1186/1477-7827-7-146. Reprod Biol Endocrinol. 2009. PMID: 20003198 Free PMC article.
-
Chromatin Accessibility and Transcriptomic Alterations in Murine Ovarian Granulosa Cells upon Deoxynivalenol Exposure.Cells. 2021 Oct 20;10(11):2818. doi: 10.3390/cells10112818. Cells. 2021. PMID: 34831041 Free PMC article.
References
-
- Simpson ER, Zhao Y, Agarwal VR, Michael MD, Bulun SE, Hinshelwood MM, Graham-Lorence S, Sun T, Fisher CR, Qin K, Mendelson CR. Aromatase expression in health and disease. Recent Prog. Horm. Res. 1997;52:185–213. - PubMed
-
- Kamat A, Hinshelwood MM, Murry BA, Mendelson CR. Mechanisms in tissue-specific regulation of estrogen biosynthesis in humans. Trends Endocrinol. Metab. 2002;13:122–128. - PubMed
-
- Kamat A, Alcorn JL, Kunczt C, Mendelson CR. Characterization of the regulatory regions of the human aromatase (P450arom) gene involved in placenta-specific expression. Mol. Endocrinol. 1998;12:1764–1777. - PubMed
-
- Jiang B, Kamat A, Mendelson CR. Hypoxia prevents induction of aromatase expression in human trophoblast cells in culture: potential inhibitory role of the hypoxia-inducible transcription factor Mash-2 (mammalian achaete-scute homologous protein-2) Mol. Endocrinol. 2000;14:1661–1673. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
