

Definition of the zebrafish genome using flow cytometry and cytogenetic mapping
- PMID: 17597531
- PMCID: PMC1925092
- DOI: 10.1186/1471-2164-8-195
Definition of the zebrafish genome using flow cytometry and cytogenetic mapping
Abstract
Background: The zebrafish (Danio rerio) is an important vertebrate model organism system for biomedical research. The syntenic conservation between the zebrafish and human genome allows one to investigate the function of human genes using the zebrafish model. To facilitate analysis of the zebrafish genome, genetic maps have been constructed and sequence annotation of a reference zebrafish genome is ongoing. However, the duplicative nature of teleost genomes, including the zebrafish, complicates accurate assembly and annotation of a representative genome sequence. Cytogenetic approaches provide "anchors" that can be integrated with accumulating genomic data.
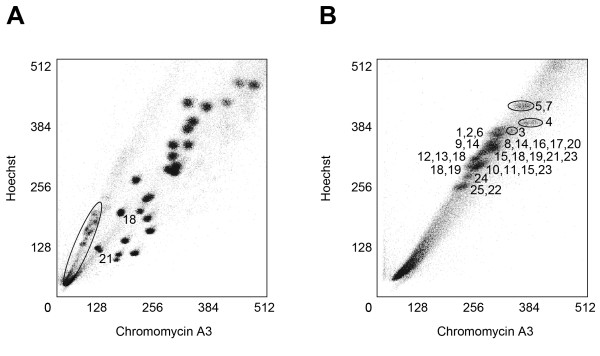
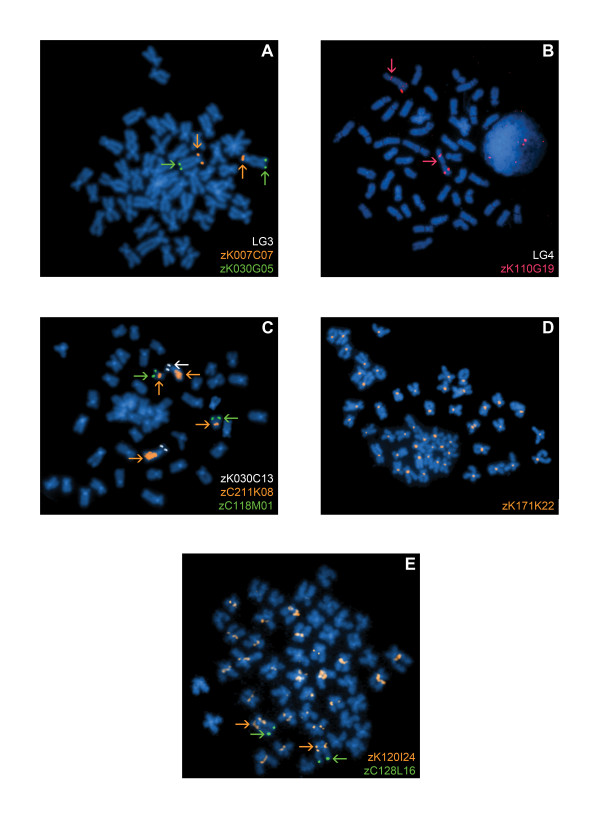
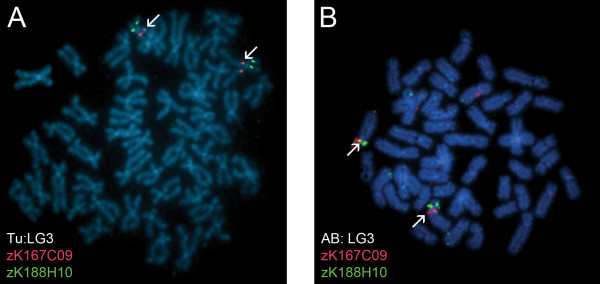
Results: Here, we cytogenetically define the zebrafish genome by first estimating the size of each linkage group (LG) chromosome using flow cytometry, followed by the cytogenetic mapping of 575 bacterial artificial chromosome (BAC) clones onto metaphase chromosomes. Of the 575 BAC clones, 544 clones localized to apparently unique chromosomal locations. 93.8% of these clones were assigned to a specific LG chromosome location using fluorescence in situ hybridization (FISH) and compared to the LG chromosome assignment reported in the zebrafish genome databases. Thirty-one BAC clones localized to multiple chromosomal locations in several different hybridization patterns. From these data, a refined second generation probe panel for each LG chromosome was also constructed.
Conclusion: The chromosomal mapping of the 575 large-insert DNA clones allows for these clones to be integrated into existing zebrafish mapping data. An accurately annotated zebrafish reference genome serves as a valuable resource for investigating the molecular basis of human diseases using zebrafish mutant models.
Figures

References
-
- Endo A, Ingalls TH. Chromosomes of the zebrafish. A model for cytogenetic, embryologic, and ecologic study. J Hered. 1968;59:382–384. - PubMed
-
- Postlethwait JH, Yan Y, Gates MA, Horne S, Amores A, Brownlie A, Donovan A, Egan ES, Force A, Gong Z, Goutel C, Fritz A, Kelsh R, Knapik E, Liao E, Paw B, Ranson D, Singer A, Thomson M, Abduljabbar TS, Yelik P, Beier D, Joly JS, Larhammar D, Rosa F, Westerfield M, Zon LI, Johnson SL, Talbot WS. Vertebrate genome evolution and the zebrafish gene map. Nat Genet. 1998;18:345–349. doi: 10.1038/ng0498-345. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
