

A complementary pair of rapid molecular screening assays for RecA activities
- PMID: 17601483
- PMCID: PMC2041836
- DOI: 10.1016/j.ab.2007.04.021
A complementary pair of rapid molecular screening assays for RecA activities
Abstract
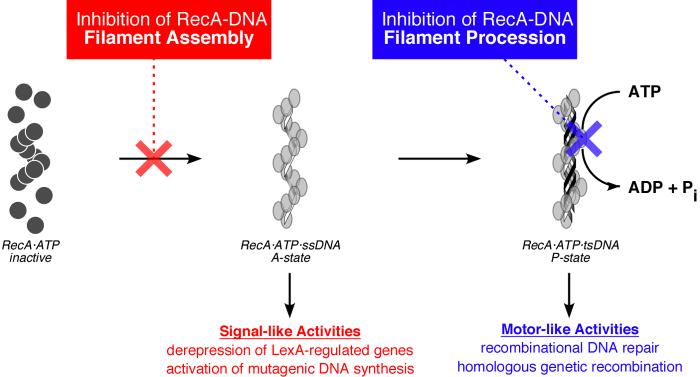
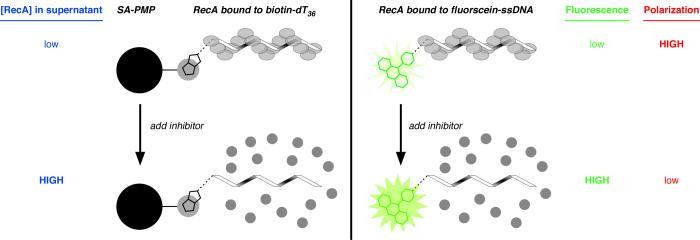
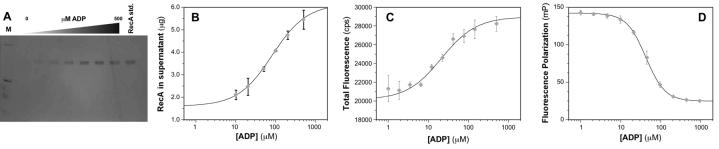
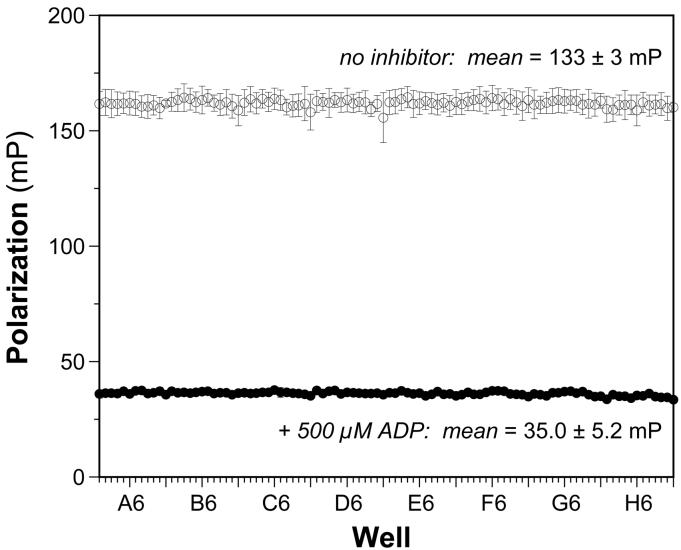
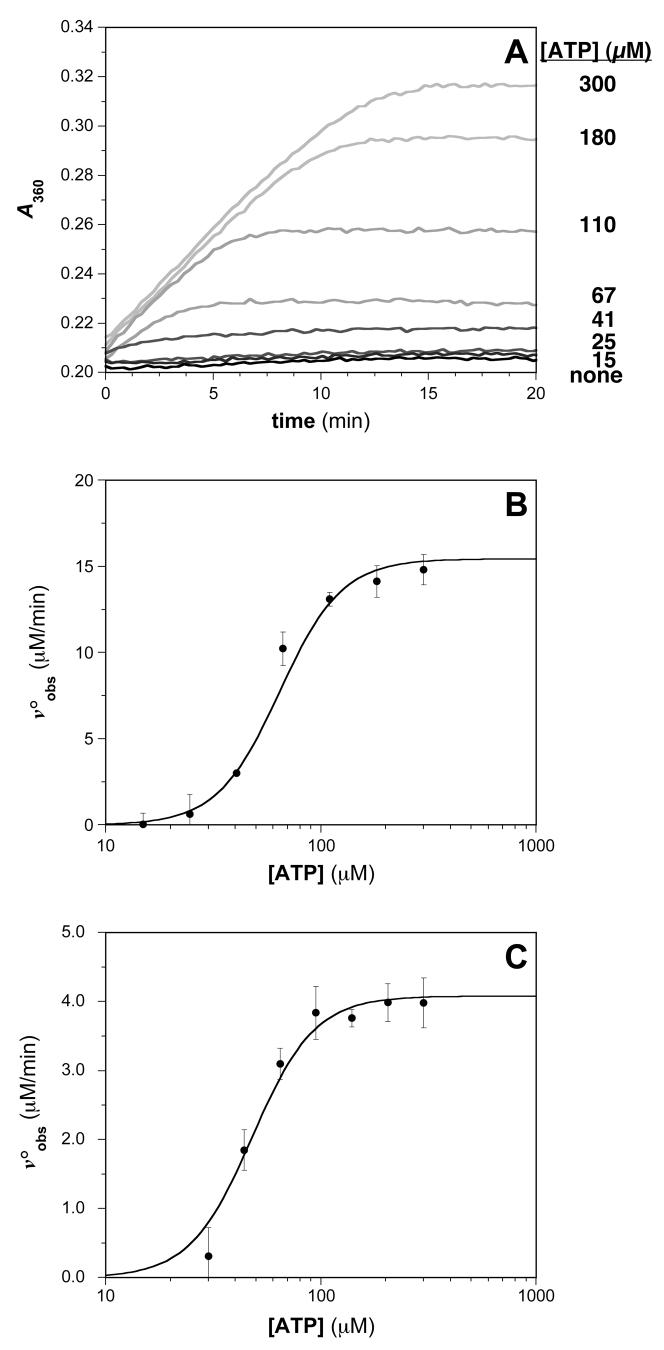
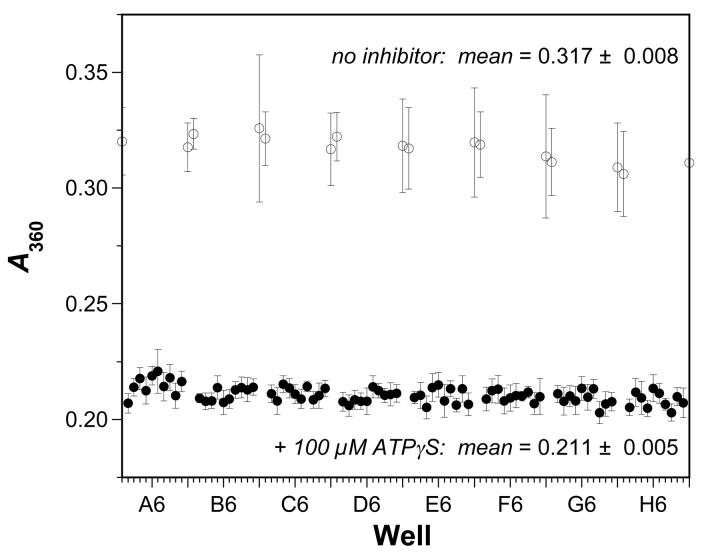
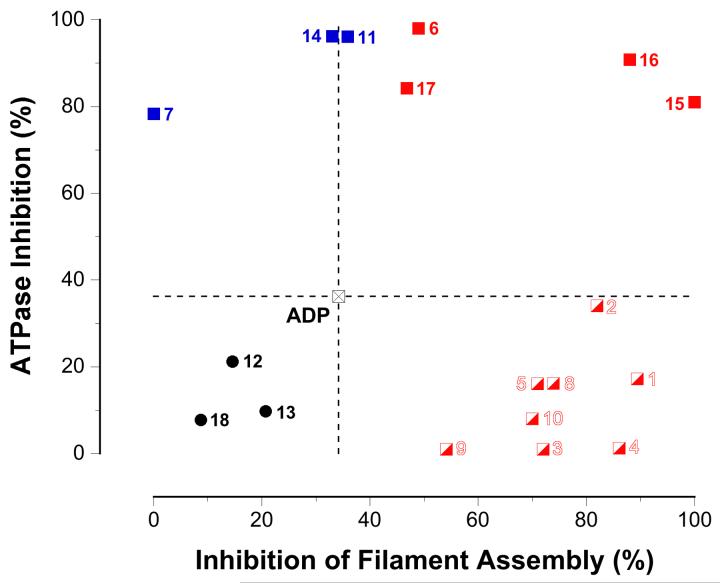
The bacterial RecA protein has been implicated in the evolution of antibiotic resistance in pathogens, which is an escalating problem worldwide. The discovery of small molecules that can selectively modulate RecA's activities can be exploited to tease apart its roles in the de novo development and transmission of antibiotic resistance genes. Toward the goal of discovering small-molecule ligands that can prevent either the assembly of an active RecA-DNA filament or its subsequent ATP-dependent motor activities, we report the design and initial validation of a pair of rapid and robust screening assays suitable for the identification of inhibitors of RecA activities. One assay is based on established methods for monitoring ATPase enzyme activity and the second is a novel assay for RecA-DNA filament assembly using fluorescence polarization. Taken together, the assay results reveal complementary sets of agents that can either suppress selectively only the ATP-driven motor activities of the RecA-DNA filament or prevent assembly of active RecA-DNA filaments altogether. The screening assays can be readily configured for use in future automated high-throughput screening projects to discover potent inhibitors that may be developed into novel adjuvants for antibiotic chemotherapy that moderate the development and transmission of antibiotic resistance genes and increase the antibiotic therapeutic index.
Figures
Similar articles
-
High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay.Assay Drug Dev Technol. 2012 Jun;10(3):260-8. doi: 10.1089/adt.2011.0409. Epub 2011 Dec 22. Assay Drug Dev Technol. 2012. PMID: 22192312 Free PMC article.
-
Directed molecular screening for RecA ATPase inhibitors.Bioorg Med Chem Lett. 2007 Jun 15;17(12):3249-53. doi: 10.1016/j.bmcl.2007.04.013. Epub 2007 Apr 10. Bioorg Med Chem Lett. 2007. PMID: 17499507 Free PMC article.
-
Inhibitors of RecA activity discovered by high-throughput screening: cell-permeable small molecules attenuate the SOS response in Escherichia coli.J Biomol Screen. 2009 Oct;14(9):1092-101. doi: 10.1177/1087057109342126. Epub 2009 Aug 12. J Biomol Screen. 2009. PMID: 19675313 Free PMC article.
-
Molecular design and functional organization of the RecA protein.Crit Rev Biochem Mol Biol. 2003;38(5):385-432. doi: 10.1080/10409230390242489. Crit Rev Biochem Mol Biol. 2003. PMID: 14693725 Review.
-
Structure and mechanism of Escherichia coli RecA ATPase.Mol Microbiol. 2005 Oct;58(2):358-66. doi: 10.1111/j.1365-2958.2005.04876.x. Mol Microbiol. 2005. PMID: 16194225 Review.
Cited by
-
Inhibitors of LexA Autoproteolysis and the Bacterial SOS Response Discovered by an Academic-Industry Partnership.ACS Infect Dis. 2018 Mar 9;4(3):349-359. doi: 10.1021/acsinfecdis.7b00122. Epub 2018 Jan 8. ACS Infect Dis. 2018. PMID: 29275629 Free PMC article.
-
Targets for Combating the Evolution of Acquired Antibiotic Resistance.Biochemistry. 2015 Jun 16;54(23):3573-82. doi: 10.1021/acs.biochem.5b00109. Epub 2015 Jun 5. Biochemistry. 2015. PMID: 26016604 Free PMC article. Review.
-
Respiration and Small Colony Variants of Staphylococcus aureus.Microbiol Spectr. 2019 May;7(3):10.1128/microbiolspec.gpp3-0069-2019. doi: 10.1128/microbiolspec.GPP3-0069-2019. Microbiol Spectr. 2019. PMID: 31198131 Free PMC article. Review.
-
Exploration of inhibitors of the bacterial LexA repressor-protease.Bioorg Med Chem Lett. 2022 Jun 1;65:128702. doi: 10.1016/j.bmcl.2022.128702. Epub 2022 Mar 26. Bioorg Med Chem Lett. 2022. PMID: 35351585 Free PMC article.
-
High-throughput screening for RecA inhibitors using a transcreener adenosine 5'-O-diphosphate assay.Assay Drug Dev Technol. 2012 Jun;10(3):260-8. doi: 10.1089/adt.2011.0409. Epub 2011 Dec 22. Assay Drug Dev Technol. 2012. PMID: 22192312 Free PMC article.
References
-
- Matic I, Taddei F, Radman M. Survival versus maintenance of genetic stability: a conflict of priorities during stress. Research in Microbiology. 2004;155:337–41. - PubMed
-
- Friedberg EC, Walker GC, Siede W. SOS responses and DNA damage tolerance in prokaryotes, DNA Repair and Mutagenesis. ASM Press; Washington, D.C.: 1995. pp. 407–464.
-
- Sassanfar M, Roberts JW. Nature of the SOS-inducing signal in Escherichia coli. The involvement of DNA replication. Journal of Molecular Biology. 1990;212:79–96. - PubMed
-
- Goodman MF. Coping with replication ‘train wrecks’ in Escherichia coli using Pol V, Pol II and RecA proteins. Trends in Biochemical Sciences. 2000;25:189–95. - PubMed
-
- Courcelle J, Ganesan AK, Hanawalt PC. Therefore, what are recombination proteins there for? Bioessays. 2001;23:463–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
