

Bacterial cyclic beta-(1,2)-glucan acts in systemic suppression of plant immune responses
- PMID: 17601826
- PMCID: PMC1955710
- DOI: 10.1105/tpc.106.047944
Bacterial cyclic beta-(1,2)-glucan acts in systemic suppression of plant immune responses
Abstract
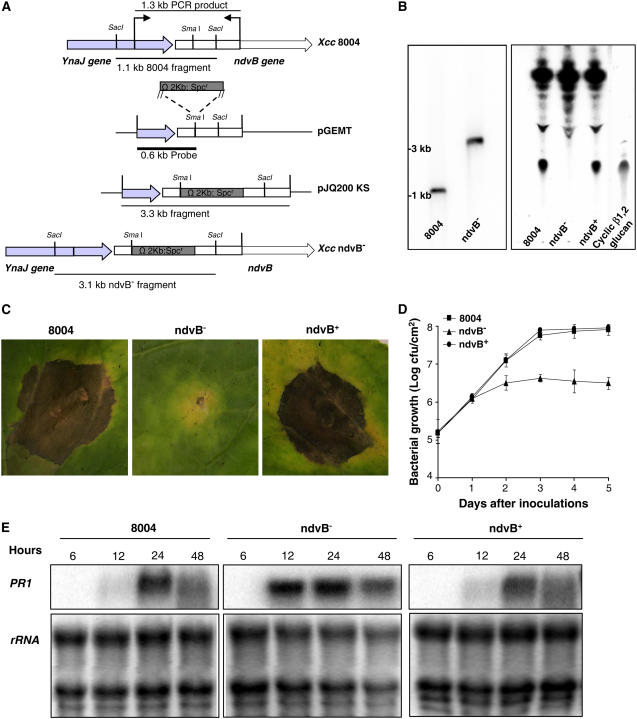
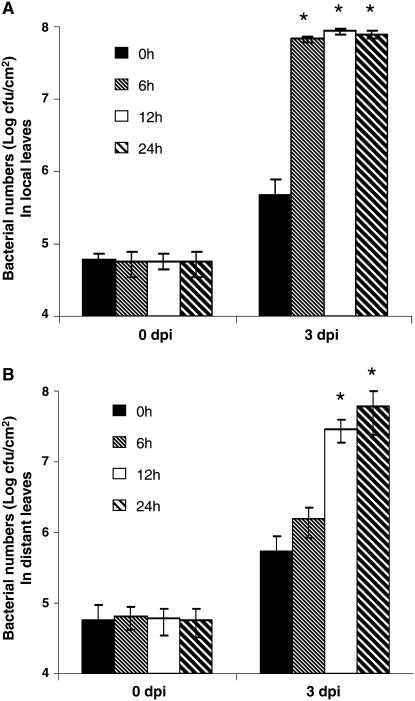
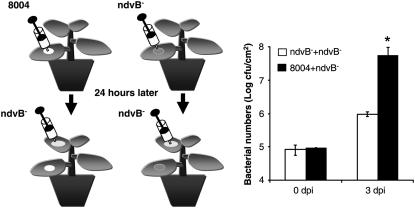
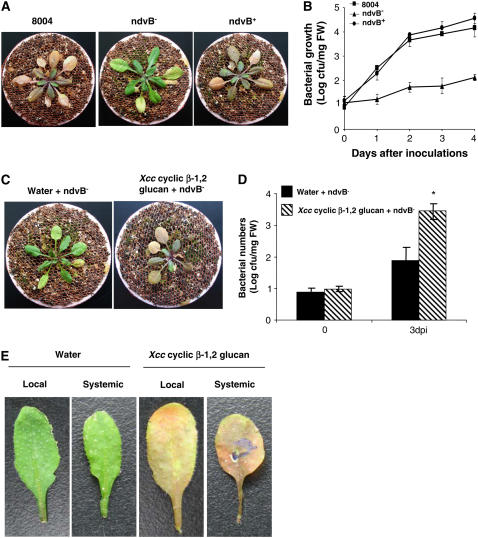
Although cyclic glucans have been shown to be important for a number of symbiotic and pathogenic bacterium-plant interactions, their precise roles are unclear. Here, we examined the role of cyclic beta-(1,2)-glucan in the virulence of the black rot pathogen Xanthomonas campestris pv campestris (Xcc). Disruption of the Xcc nodule development B (ndvB) gene, which encodes a glycosyltransferase required for cyclic glucan synthesis, generated a mutant that failed to synthesize extracellular cyclic beta-(1,2)-glucan and was compromised in virulence in the model plants Arabidopsis thaliana and Nicotiana benthamiana. Infection of the mutant bacterium in N. benthamiana was associated with enhanced callose deposition and earlier expression of the PATHOGENESIS-RELATED1 (PR-1) gene. Application of purified cyclic beta-(1,2)-glucan prior to inoculation of the ndvB mutant suppressed the accumulation of callose deposition and the expression of PR-1 in N. benthamiana and restored virulence in both N. benthamiana and Arabidopsis plants. These effects were seen when cyclic glucan and bacteria were applied either to the same or to different leaves. Cyclic beta-(1,2)-glucan-induced systemic suppression was associated with the transport of the molecule throughout the plant. Systemic suppression is a novel counterdefensive strategy that may facilitate pathogen spread in plants and may have important implications for the understanding of plant-pathogen coevolution and for the development of phytoprotection measures.
Figures




References
-
- Abramovitch, R.B., and Martin, G.B. (2004). Strategies used by bacterial pathogens to suppress plant defenses. Curr. Opin. Plant Biol. 7 356–364. - PubMed
-
- Arellano-Reynoso, B., et al. (2005). Cyclic beta-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nat. Immunol. 6 618–625. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
