

Recognition of the inner lipoyl-bearing domain of dihydrolipoyl transacetylase and of the blood glucose-lowering compound AZD7545 by pyruvate dehydrogenase kinase 2
- PMID: 17602666
- PMCID: PMC2136408
- DOI: 10.1021/bi700650k
Recognition of the inner lipoyl-bearing domain of dihydrolipoyl transacetylase and of the blood glucose-lowering compound AZD7545 by pyruvate dehydrogenase kinase 2
Abstract
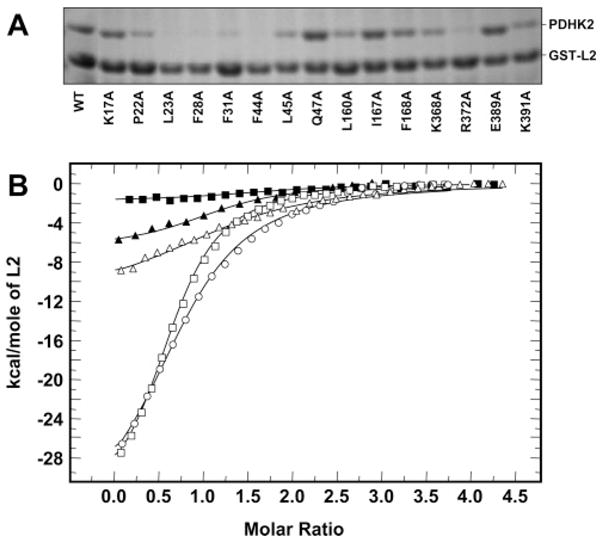
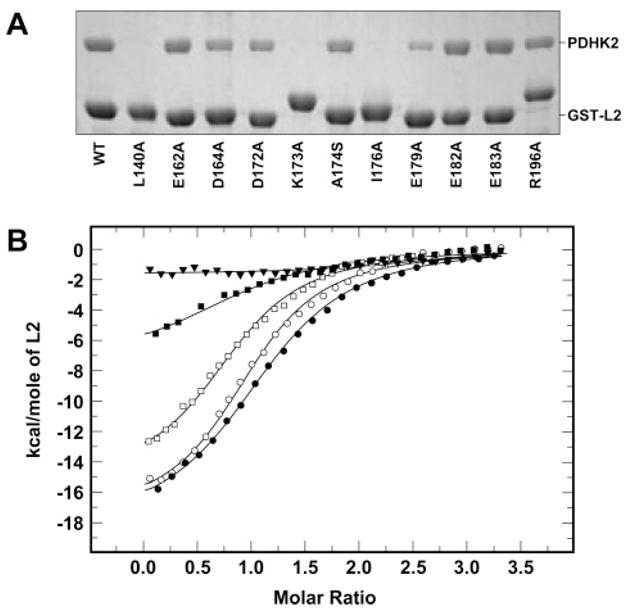
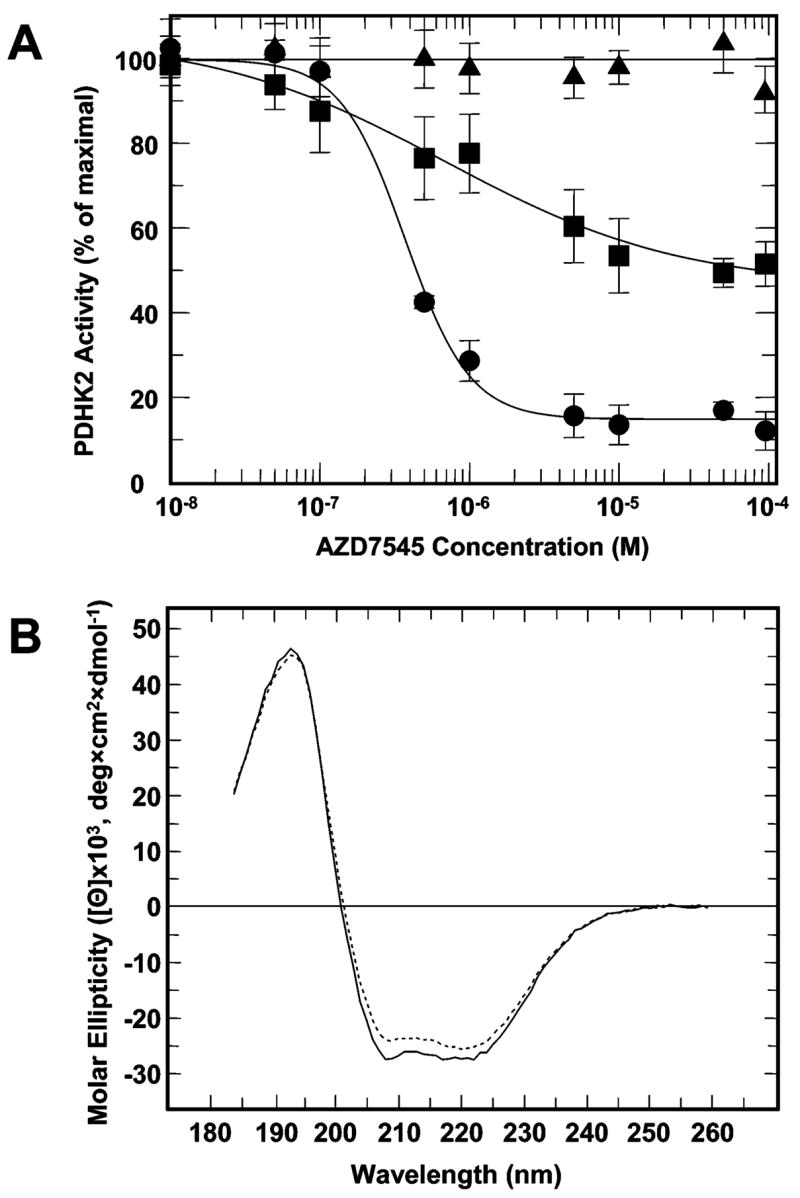
Pyruvate dehydrogenase kinase 2 (PDHK2) is a unique mitochondrial protein kinase that regulates the activity of the pyruvate dehydrogenase multienzyme complex (PDC). PDHK2 is an integral component of PDC tightly bound to the inner lipoyl-bearing domains (L2) of the dihydrolipoyl transacetylase component (E2) of PDC. This association has been reported to bring about an up to 10-fold increase in kinase activity. Despite the central role played by E2 in the maintenance of PDHK2 functionality in the PDC-bound state, the molecular mechanisms responsible for the recognition of L2 by PDHK2 and for the E2-dependent PDHK2 activation are largely unknown. In this study, we used a combination of molecular modeling and site-directed mutagenesis to identify the amino acid residues essential for the interaction between PDHK2 and L2 and for the activation of PDHK2 by E2. On the basis of the results of site-directed mutagenesis, it appears that a number of PDHK2 residues located in its R domain (P22, L23, F28, F31, F44, L45, and L160) and in the so-called "cross arm" structure (K368, R372, and K391) are critical in determining the strength of the interaction between PDHK2 and L2. The residues of L2 essential for recognition by PDHK2 include L140, K173, I176, E179, and to a lesser extent D164, D172, and A174. Importantly, certain PDHK2 residues forming interfaces with L2, i.e., K17, P22, F31, F44, R372, and K391, are also critical for the maintenance of enhanced PDHK2 activity in the E2-bound state. Finally, evidence that the blood glucose-lowering compound AZD7545 disrupts the interactions between PDHK2 and L2 and thereby inhibits PDHK2 activity is presented.
Figures


Similar articles
-
Specific ion influences on self-association of pyruvate dehydrogenase kinase isoform 2 (PDHK2), binding of PDHK2 to the L2 lipoyl domain, and effects of the lipoyl group-binding site inhibitor, Nov3r.Biochemistry. 2008 Feb 26;47(8):2312-24. doi: 10.1021/bi7014772. Epub 2008 Jan 26. Biochemistry. 2008. PMID: 18220415
-
Structural and functional insights into the molecular mechanisms responsible for the regulation of pyruvate dehydrogenase kinase 2.J Biol Chem. 2008 Jun 6;283(23):15789-98. doi: 10.1074/jbc.M800311200. Epub 2008 Apr 3. J Biol Chem. 2008. PMID: 18387944 Free PMC article.
-
Allosteric coupling in pyruvate dehydrogenase kinase 2.Biochemistry. 2008 Aug 12;47(32):8358-66. doi: 10.1021/bi800631h. Epub 2008 Jul 16. Biochemistry. 2008. PMID: 18627174 Free PMC article.
-
Pyruvate dehydrogenase kinase regulatory mechanisms and inhibition in treating diabetes, heart ischemia, and cancer.Cell Mol Life Sci. 2007 Apr;64(7-8):830-49. doi: 10.1007/s00018-007-6380-z. Cell Mol Life Sci. 2007. PMID: 17310282 Free PMC article. Review.
-
Distinct regulatory properties of pyruvate dehydrogenase kinase and phosphatase isoforms.Prog Nucleic Acid Res Mol Biol. 2001;70:33-75. doi: 10.1016/s0079-6603(01)70013-x. Prog Nucleic Acid Res Mol Biol. 2001. PMID: 11642366 Review.
Cited by
-
Structure-guided development of specific pyruvate dehydrogenase kinase inhibitors targeting the ATP-binding pocket.J Biol Chem. 2014 Feb 14;289(7):4432-43. doi: 10.1074/jbc.M113.533885. Epub 2013 Dec 19. J Biol Chem. 2014. PMID: 24356970 Free PMC article.
-
Elucidation of the interaction loci of the human pyruvate dehydrogenase complex E2·E3BP core with pyruvate dehydrogenase kinase 1 and kinase 2 by H/D exchange mass spectrometry and nuclear magnetic resonance.Biochemistry. 2015 Jan 13;54(1):69-82. doi: 10.1021/bi5013113. Epub 2014 Dec 17. Biochemistry. 2015. PMID: 25436986 Free PMC article.
-
Distinct structural mechanisms for inhibition of pyruvate dehydrogenase kinase isoforms by AZD7545, dichloroacetate, and radicicol.Structure. 2007 Aug;15(8):992-1004. doi: 10.1016/j.str.2007.07.001. Epub 2007 Aug 2. Structure. 2007. PMID: 17683942 Free PMC article.
-
Global view of cognate kinase activation by the human pyruvate dehydrogenase complex.Sci Rep. 2017 Feb 23;7:42760. doi: 10.1038/srep42760. Sci Rep. 2017. PMID: 28230160 Free PMC article.
-
Pyruvate dehydrogenase kinase-4 structures reveal a metastable open conformation fostering robust core-free basal activity.J Biol Chem. 2008 Sep 12;283(37):25305-25315. doi: 10.1074/jbc.M802249200. Epub 2008 Jul 24. J Biol Chem. 2008. PMID: 18658136 Free PMC article.
References
-
- Randle PJ. Metabolic fuel selection: General integration at the whole-body level. Proc Nutr Soc. 1995;54:317–327. - PubMed
-
- Holness MJ, Sugden MC. Regulation of pyruvate dehydrogenase complex activity by reversible phosphorylation. Biochem Soc Trans. 2003;31:1143–1151. - PubMed
-
- Patel MS, Roche TE. Molecular biology and biochemistry of pyruvate dehydrogenase complexes. FASEB J. 1990;4:3224–3233. - PubMed
-
- Ravindran S, Radke GA, Guest JR, Roche TE. Lipoyl domain-based mechanism for the integrated feedback control of the pyruvate dehydrogenase complex by enhancement of pyruvate dehydrogenase kinase activity. J Biol Chem. 1996;271:653–662. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
