

A mutation in EXO1 defines separable roles in DNA mismatch repair and post-replication repair
- PMID: 17602897
- PMCID: PMC2447855
- DOI: 10.1016/j.dnarep.2007.05.004
A mutation in EXO1 defines separable roles in DNA mismatch repair and post-replication repair
Abstract
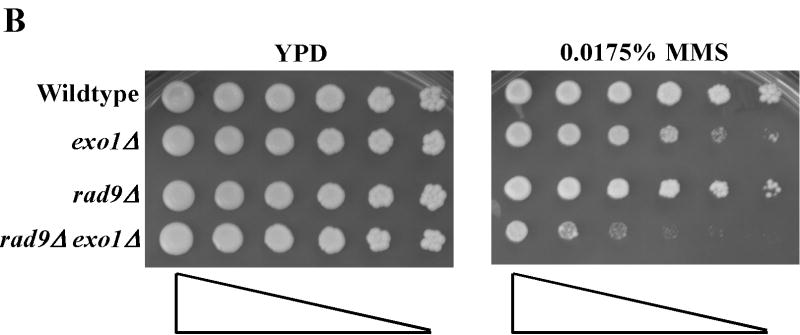
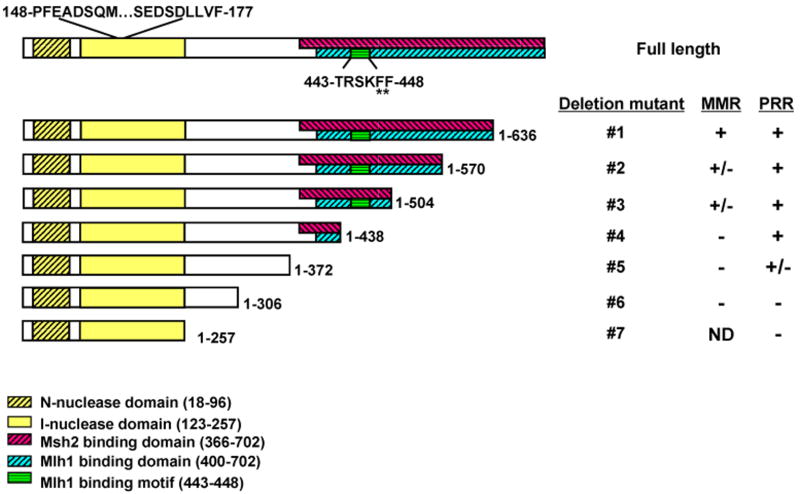
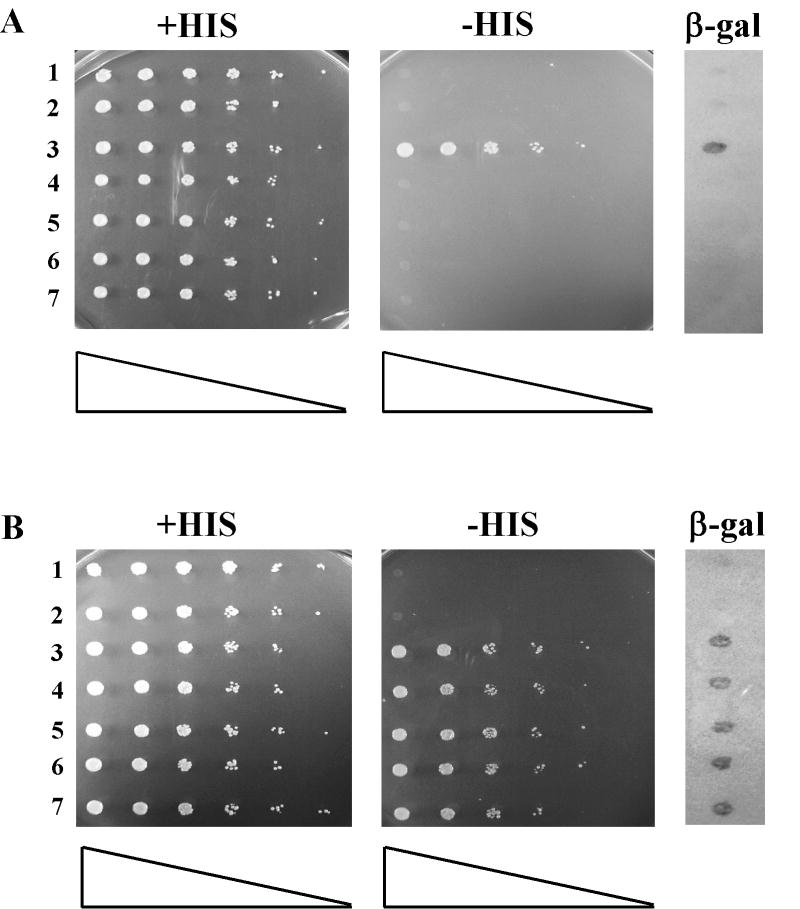
Replication forks stall at DNA lesions or as a result of an unfavorable replicative environment. These fork stalling events have been associated with recombination and gross chromosomal rearrangements. Recombination and fork bypass pathways are the mechanisms accountable for restart of stalled forks. An important lesion bypass mechanism is the highly conserved post-replication repair (PRR) pathway that is composed of error-prone translesion and error-free bypass branches. EXO1 codes for a Rad2p family member nuclease that has been implicated in a multitude of eukaryotic DNA metabolic pathways that include DNA repair, recombination, replication, and telomere integrity. In this report, we show EXO1 functions in the MMS2 error-free branch of the PRR pathway independent of the role of EXO1 in DNA mismatch repair (MMR). Consistent with the idea that EXO1 functions independently in two separate pathways, we defined a domain of Exo1p required for PRR distinct from those required for interaction with MMR proteins. We then generated a point mutant exo1 allele that was defective for the function of Exo1p in MMR due to disrupted interaction with Mlh1p, but still functional for PRR. Lastly, by using a compound exo1 mutant that was defective for interaction with Mlh1p and deficient for nuclease activity, we provide further evidence that Exo1p plays both structural and catalytic roles during MMR.
Figures






References
-
- Branzei D, Foiani M. The DNA damage response during DNA replication. Curr Opin Cell Biol. 2005;17:568–575. - PubMed
-
- Waga S, Stillman B. The DNA replication fork in eukaryotic cells. Annu Rev Biochem. 1998;67:721–751. - PubMed
-
- Kolodner RD, Putnam CD, Myung K. Maintenance of genome stability in Saccharomyces cerevisiae. Science. 2002;297:552–557. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, Guldberg P, Sehested M, Nesland JM, Lukas C, Orntoft T, Lukas J, Bartek J. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
-
- Gorgoulis VG, Vassiliou LV, Karakaidos P, Zacharatos P, Kotsinas A, Liloglou T, Venere M, Ditullio RA, Jr, Kastrinakis NG, Levy B, Kletsas D, Yoneta A, Herlyn M, Kittas C, Halazonetis TD. Activation of the DNA damage checkpoint and genomic instability in human precancerous lesions. Nature. 2005;434:907–913. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
