

Early events in the evolution of the Silene latifolia Y chromosome: male specialization and recombination arrest
- PMID: 17603119
- PMCID: PMC2013713
- DOI: 10.1534/genetics.107.071175
Early events in the evolution of the Silene latifolia Y chromosome: male specialization and recombination arrest
Abstract
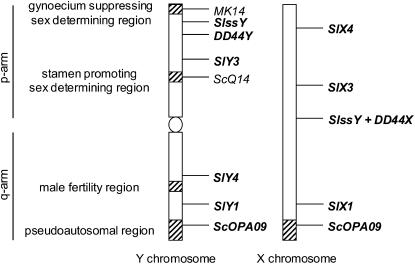
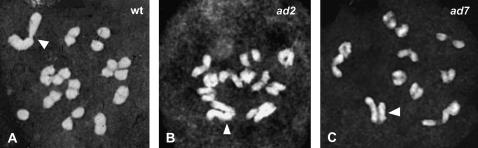
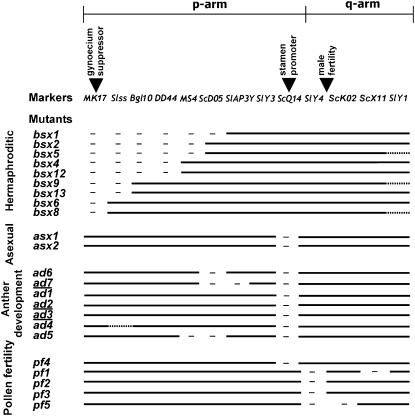
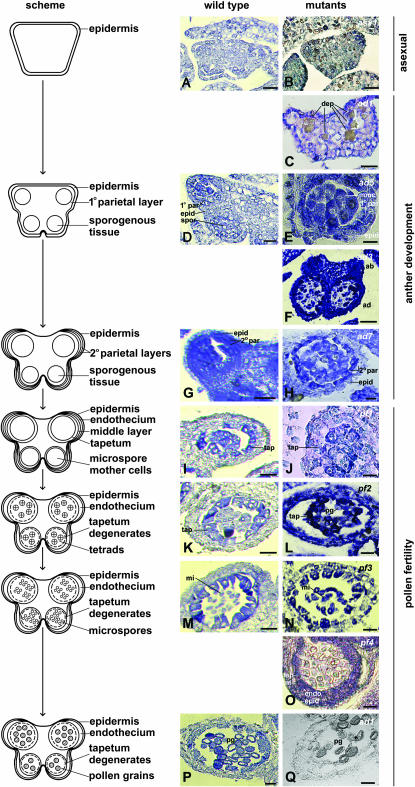
Understanding the origin and evolution of sex chromosomes requires studying recently evolved X-Y chromosome systems such as those in some flowering plants. We describe Y chromosome deletion mutants of Silene latifolia, a dioecious plant with heteromorphic sex chromosomes. The combination of results from new and previously described deletions with histological descriptions of their stamen development defects indicates the presence of two distinct Y regions containing loci with indispensable roles in male reproduction. We determined their positions relative to the two main sex determination functions (female suppressing and the other male promoting). A region proximal to the centromere on the Y p arm containing the putative stamen promoting sex determination locus includes additional early stamen developmental factors. A medial region of the Y q arm carries late pollen fertility factors. Cytological analysis of meiotic X-Y pairing in one of the male-sterile mutants indicates that the Y carries sequences or functions specifically affecting sex chromosome pairing.
Figures
Similar articles
-
The inter-specific hybrid Silene latifolia x S. viscosa reveals early events of sex chromosome evolution.Evol Dev. 2005 Jul-Aug;7(4):327-36. doi: 10.1111/j.1525-142X.2005.05038.x. Evol Dev. 2005. PMID: 15982369
-
DNA diversity in sex-linked and autosomal genes of the plant species Silene latifolia and Silene dioica.Mol Biol Evol. 2001 Aug;18(8):1442-54. doi: 10.1093/oxfordjournals.molbev.a003930. Mol Biol Evol. 2001. PMID: 11470835
-
An accumulation of tandem DNA repeats on the Y chromosome in Silene latifolia during early stages of sex chromosome evolution.Chromosoma. 2006 Oct;115(5):376-82. doi: 10.1007/s00412-006-0065-5. Epub 2006 Apr 13. Chromosoma. 2006. PMID: 16612641
-
The genomics of plant sex chromosomes.Plant Sci. 2015 Jul;236:126-35. doi: 10.1016/j.plantsci.2015.03.019. Epub 2015 Apr 2. Plant Sci. 2015. PMID: 26025526 Review.
-
A cytogenetic view of sex chromosome evolution in plants.Cytogenet Genome Res. 2008;120(3-4):241-6. doi: 10.1159/000121073. Epub 2008 May 23. Cytogenet Genome Res. 2008. PMID: 18504353 Review.
Cited by
-
The Silene latifolia genome and its giant Y chromosome.Science. 2025 Feb 7;387(6734):630-636. doi: 10.1126/science.adj7430. Epub 2025 Feb 6. Science. 2025. PMID: 39913565
-
Defining regions and rearrangements of the Silene latifolia Y chromosome.Genetics. 2008 Apr;178(4):2045-53. doi: 10.1534/genetics.107.084566. Epub 2008 Feb 3. Genetics. 2008. PMID: 18245827 Free PMC article.
-
A new plant sex-linked gene with high sequence diversity and possible introgression of the X copy.Heredity (Edinb). 2011 Feb;106(2):339-47. doi: 10.1038/hdy.2010.76. Epub 2010 Jun 16. Heredity (Edinb). 2011. PMID: 20551975 Free PMC article.
-
Sexy ways: approaches to studying plant sex chromosomes.J Exp Bot. 2024 Sep 11;75(17):5204-5219. doi: 10.1093/jxb/erae173. J Exp Bot. 2024. PMID: 38652048 Free PMC article. Review.
-
Independent origin of sex chromosomes in two species of the genus Silene.Genetics. 2008 Jun;179(2):1129-33. doi: 10.1534/genetics.107.085670. Genetics. 2008. PMID: 18558658 Free PMC article.
References
-
- Ainsworth, C., J. Parker and V. Buchanan-Wollaston, 1998. Sex determination in plants. Curr. Top. Dev. Biol. 38: 167–223. - PubMed
-
- Atanassov, I, C. Delichere, D. A. Filatov, D. Charlesworth, I. Negrutiu et al., 2001. Analysis and evolution of two functional Y-linked loci in a plant sex chromosome system. Mol. Biol. Evol. 18: 2162–2168. - PubMed
-
- Brooks, R., 2000. Negative genetic correlation between male sexual attractiveness and survival. Nature 406: 67–70. - PubMed
-
- Bull, J. J., 1983. Evolution of Sex Determining Mechanisms. Benjamin/Cummings Publishing Company, Menlo Park, CA.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
