

Molecular motor-induced instabilities and cross linkers determine biopolymer organization
- PMID: 17604319
- PMCID: PMC2098725
- DOI: 10.1529/biophysj.106.095919
Molecular motor-induced instabilities and cross linkers determine biopolymer organization
Abstract
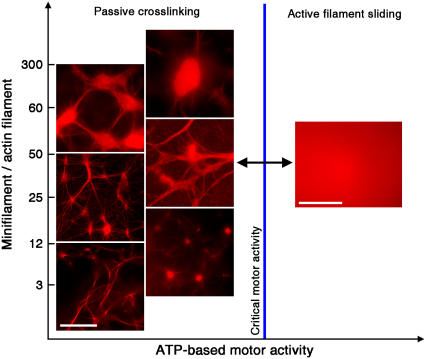
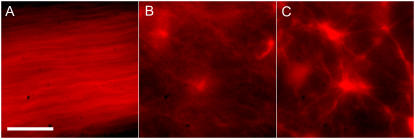
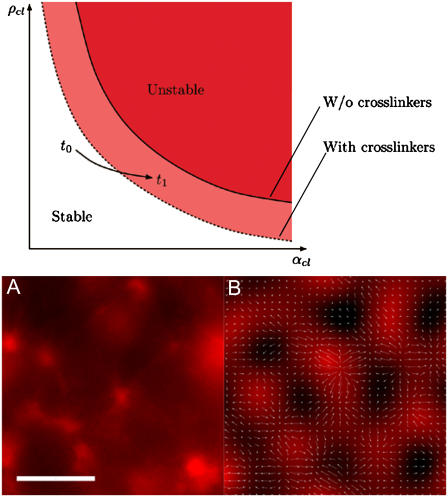
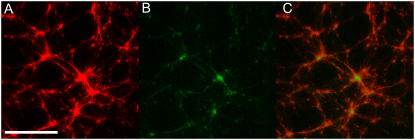
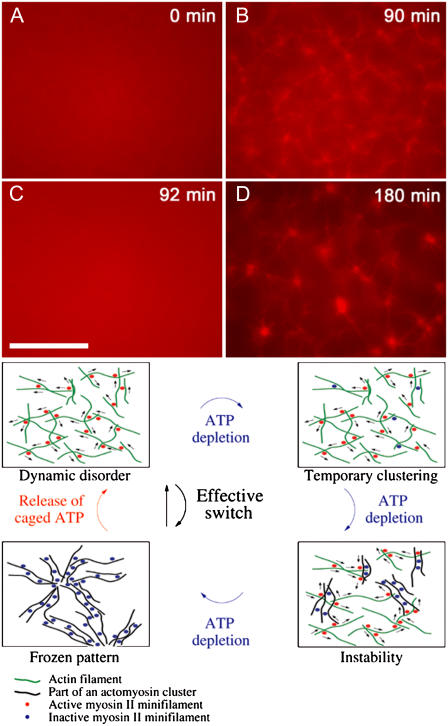
All eukaryotic cells rely on the active self-organization of protein filaments to form a responsive intracellular cytoskeleton. The necessity of motility and reaction to stimuli additionally requires pathways that quickly and reversibly change cytoskeletal organization. While thermally driven order-disorder transitions are, from the viewpoint of physics, the most obvious method for controlling states of organization, the timescales necessary for effective cellular dynamics would require temperatures exceeding the physiologically viable temperature range. We report a mechanism whereby the molecular motor myosin II can cause near-instantaneous order-disorder transitions in reconstituted cytoskeletal actin solutions. When motor-induced filament sliding diminishes, the actin network structure rapidly and reversibly self-organizes into various assemblies. Addition of stable cross linkers was found to alter the architectures of ordered assemblies. These isothermal transitions between dynamic disorder and self-assembled ordered states illustrate that the interplay between passive crosslinking and molecular motor activity plays a substantial role in dynamic cellular organization.
Figures

References
-
- Borisy, G. G., and T. M. Svitkina. 2000. Actin machinery: pushing the envelope. Curr. Opin. Cell Biol. 12:104–112. - PubMed
-
- Pollard, T. D. 2001. Genomics, the cytoskeleton and motility. Nature. 409:842–843. - PubMed
-
- Pantaloni, D., C. Le Clainche, and M. F. Carlier. 2001. Mechanism of actin-based motility. Science. 292:1502–1506. - PubMed
-
- Bershadsky, A. D., N. Q. Balaban, and B. Geiger. 2003. Adhesion-dependent cell mechanosensitivity. Annu. Rev. Cell Dev. Biol. 19:677–695. - PubMed
-
- Ingber, D. E. 2002. Mechanical signaling. Ann. N. Y. Acad. Sci. 961:162–163. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
