

Restoration of calbindin after fetal hippocampal CA3 cell grafting into the injured hippocampus in a rat model of temporal lobe epilepsy
- PMID: 17604349
- PMCID: PMC3612498
- DOI: 10.1002/hipo.20311
Restoration of calbindin after fetal hippocampal CA3 cell grafting into the injured hippocampus in a rat model of temporal lobe epilepsy
Abstract
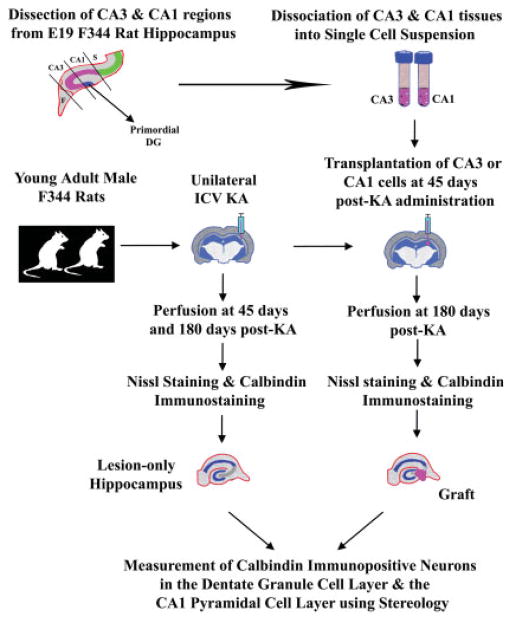
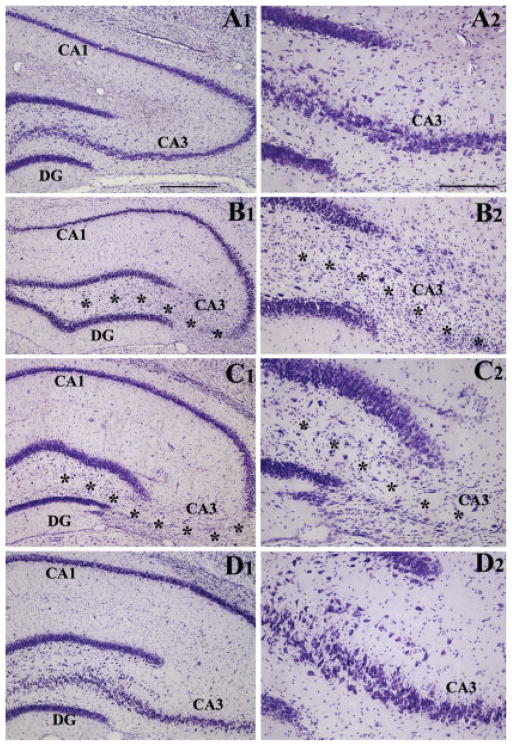
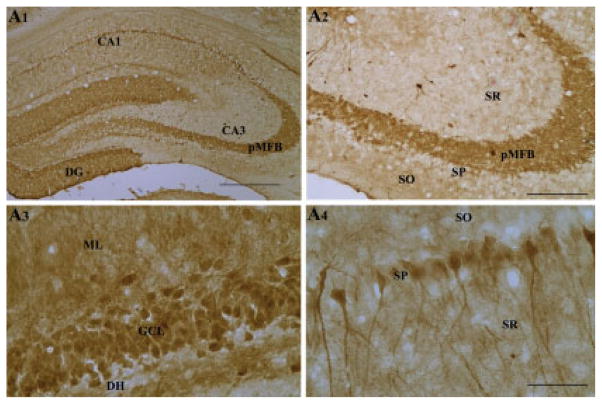
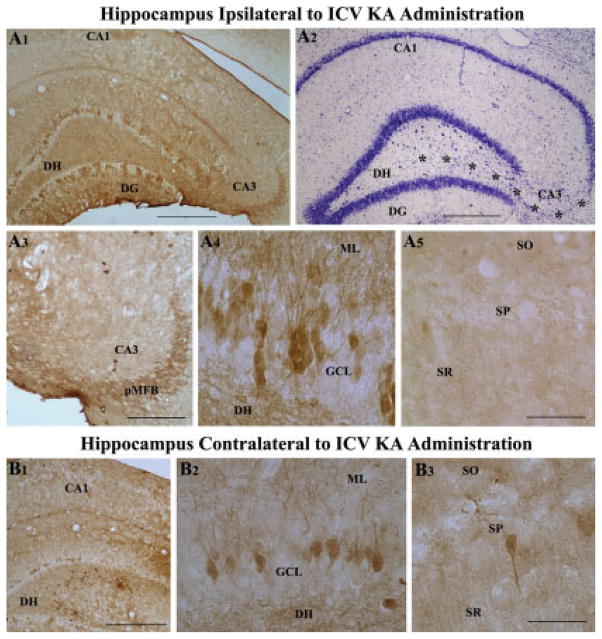
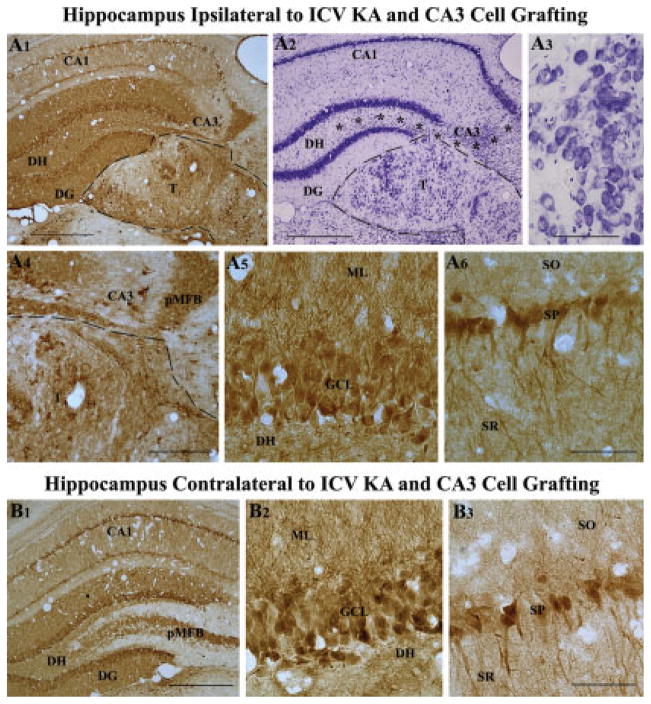
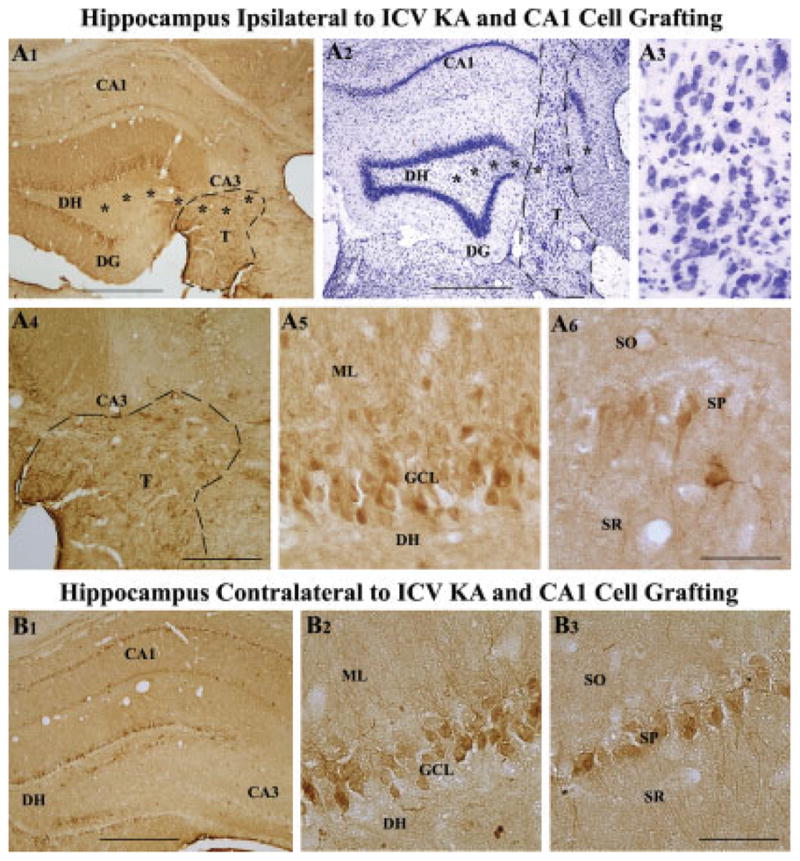
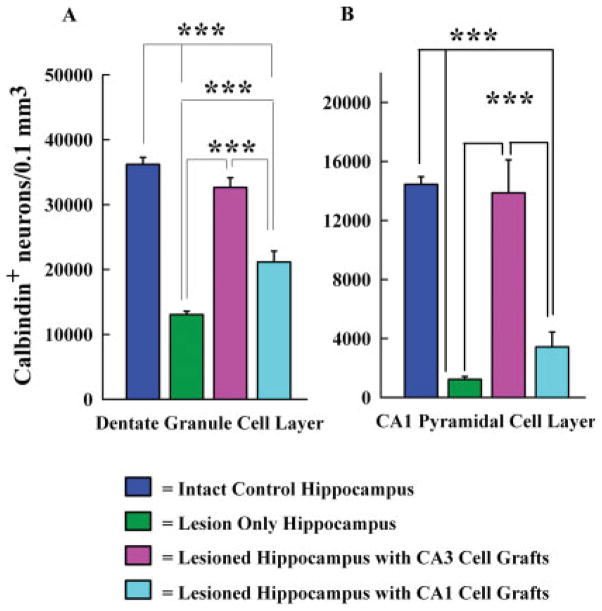
Degeneration of the CA3 pyramidal and dentate hilar neurons in the adult rat hippocampus after an intracerebroventricular kainic acid (KA) administration, a model of temporal lobe epilepsy, leads to permanent loss of the calcium binding protein calbindin in major fractions of dentate granule cells and CA1 pyramidal neurons. We hypothesize that the enduring loss of calbindin in the dentate gyrus and the CA1 subfield after CA3-lesion is due to disruption of the hippocampal circuitry leading to hyperexcitability in these regions; therefore, specific cell grafts that are capable of both reconstructing the disrupted circuitry and suppressing hyperexcitability in the injured hippocampus can restore calbindin. We compared the effects of fetal CA3 or CA1 cell grafting into the injured CA3 region of adult rats at 45 days after KA-induced injury on the hippocampal calbindin. The calbindin immunoreactivity in the dentate granule cells and the CA1 pyramidal neurons of grafted animals was evaluated at 6 months after injury (i.e. at 4.5 months post-grafting). Compared with the intact hippocampus, the calbindin in "lesion-only" hippocampus was dramatically reduced at 6 months post-lesion. However, calbindin expression was restored in the lesioned hippocampus receiving CA3 cell grafts. In contrast, in the lesioned hippocampus receiving CA1 cell grafts, calbindin expression remained less than the intact hippocampus. Thus, specific cell grafting restores the injury-induced loss of calbindin in the adult hippocampus, likely via restitution of the disrupted circuitry. Since loss of calbindin after hippocampal injury is linked to hyperexcitability, re-expression of calbindin in both dentate gyrus and CA1 subfield following CA3 cell grafting may suggest that specific cell grafting is efficacious for ameliorating injury-induced hyperexcitability in the adult hippocampus. However, electrophysiological studies of KA-lesioned hippocampus receiving CA3 cell grafts are required in future to validate this possibility.
(c) 2007 Wiley-Liss, Inc.
Figures
References
-
- Baimbridge KG. Calcium-binding proteins in the dentate gyrus. Epilepsy Res Suppl. 1992;7:211–220. - PubMed
-
- Baimbridge KG, Miller JJ. Immunohistochemical localization of calcium-binding protein in the cerebellum, hippocampal formation and olfactory bulb of the rat. Brain Res. 1982;245:223–229. - PubMed
-
- Baimbridge KG, Miller JJ. Hippocampal calcium-binding protein during commissural kindling-induced epileptogenesis: Progressive decline and effects of anticonvulsants. Brain Res. 1984;324:85–90. - PubMed
-
- Baimbridge KG, Mody I, Miller JJ. Reduction of rat hippocampal calcium-binding protein following commissural, amygdala, septal, perforant path, and olfactory bulb kindling. Epilepsia. 1985;26:460–465. - PubMed
-
- Baimbridge KG, Celio MR, Rogers JH. Calcium-binding proteins in the nervous system. Trends Neurosci. 1992;15:303–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
