

Genome-wide fitness test and mechanism-of-action studies of inhibitory compounds in Candida albicans
- PMID: 17604452
- PMCID: PMC1904411
- DOI: 10.1371/journal.ppat.0030092
Genome-wide fitness test and mechanism-of-action studies of inhibitory compounds in Candida albicans
Abstract
Candida albicans is a prevalent fungal pathogen amongst the immunocompromised population, causing both superficial and life-threatening infections. Since C. albicans is diploid, classical transmission genetics can not be performed to study specific aspects of its biology and pathogenesis. Here, we exploit the diploid status of C. albicans by constructing a library of 2,868 heterozygous deletion mutants and screening this collection using 35 known or novel compounds to survey chemically induced haploinsufficiency in the pathogen. In this reverse genetic assay termed the fitness test, genes related to the mechanism of action of the probe compounds are clearly identified, supporting their functional roles and genetic interactions. In this report, chemical-genetic relationships are provided for multiple FDA-approved antifungal drugs (fluconazole, voriconazole, caspofungin, 5-fluorocytosine, and amphotericin B) as well as additional compounds targeting ergosterol, fatty acid and sphingolipid biosynthesis, microtubules, actin, secretion, rRNA processing, translation, glycosylation, and protein folding mechanisms. We also demonstrate how chemically induced haploinsufficiency profiles can be used to identify the mechanism of action of novel antifungal agents, thereby illustrating the potential utility of this approach to antifungal drug discovery.
Conflict of interest statement
Figures
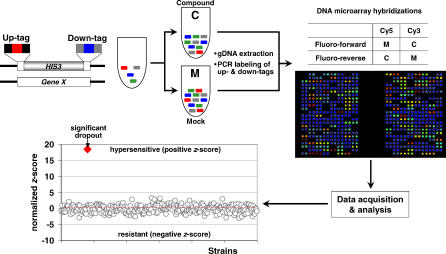
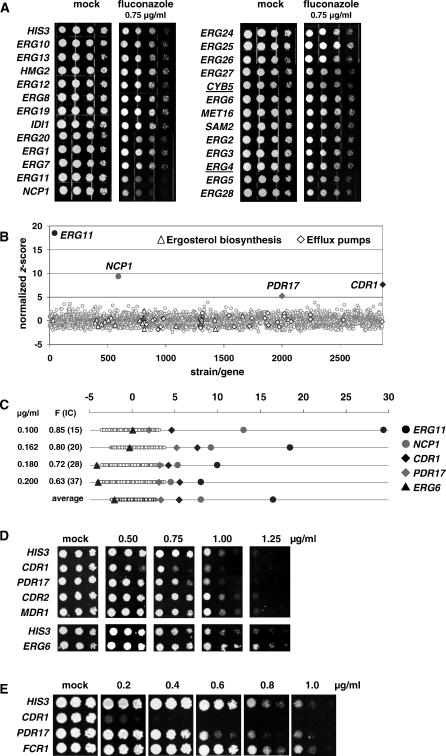
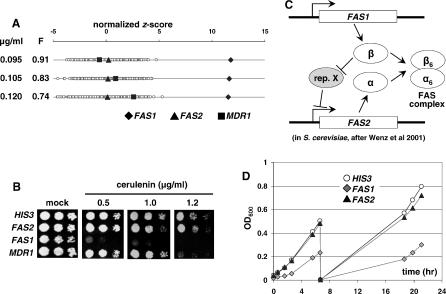
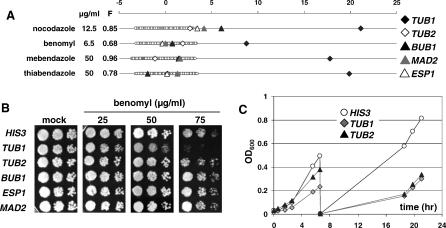
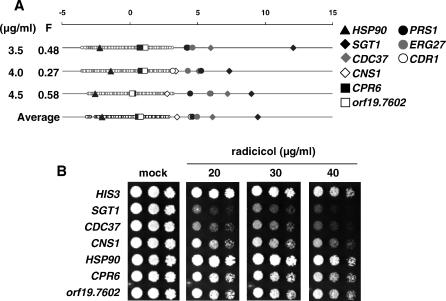
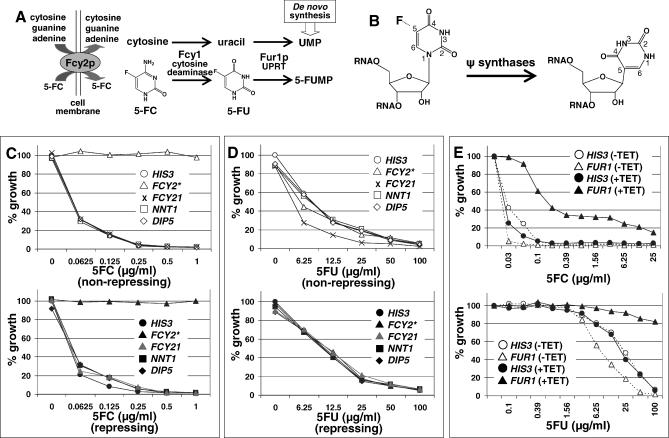
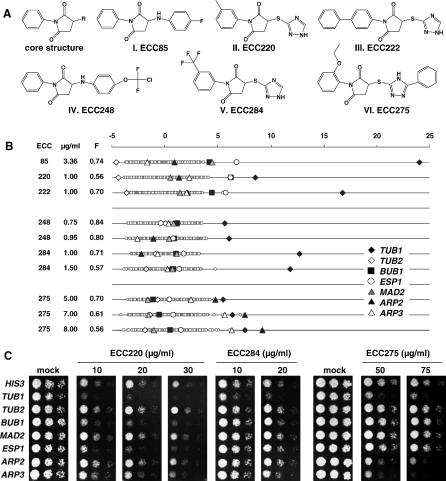
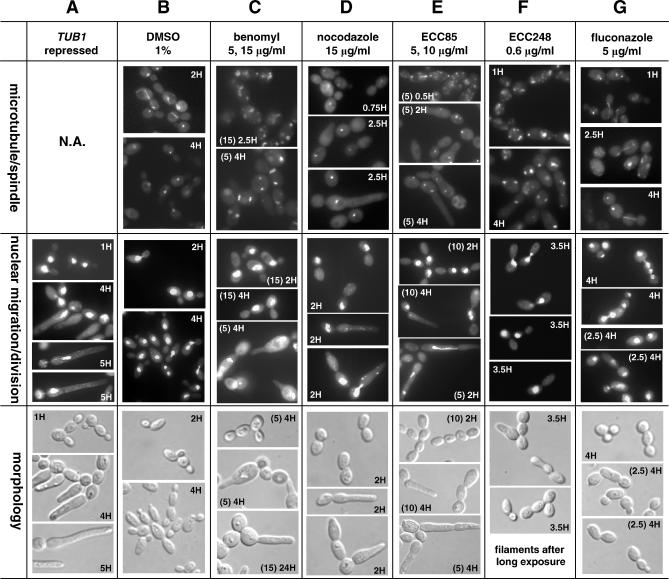
References
-
- Pappas PG, Rex JH, Lee J, Hamill RJ, Larsen RA, et al. A prospective observational study of candidemia: Epidemiology, therapy, and influences on mortality in hospitalized adult and pediatric patients. Clin Infect Dis. 2003;37:634–643. - PubMed
-
- Wisplinghoff H, Bischoff T, Tallent SM, Seifert H, Wenzel RP, et al. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin Infect Dis. 2004;39:309–317. - PubMed
-
- Kellis M, Birren BW, Lander ES. Proof and evolutionary analysis of ancient genome duplication in the yeast Saccharomyces cerevisiae . Nature. 2004;428:617–624. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
