

The evolution of N-glycan-dependent endoplasmic reticulum quality control factors for glycoprotein folding and degradation
- PMID: 17606910
- PMCID: PMC1905923
- DOI: 10.1073/pnas.0704862104
The evolution of N-glycan-dependent endoplasmic reticulum quality control factors for glycoprotein folding and degradation
Abstract
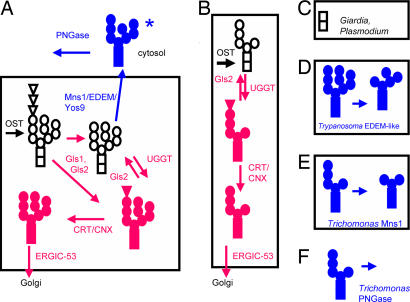
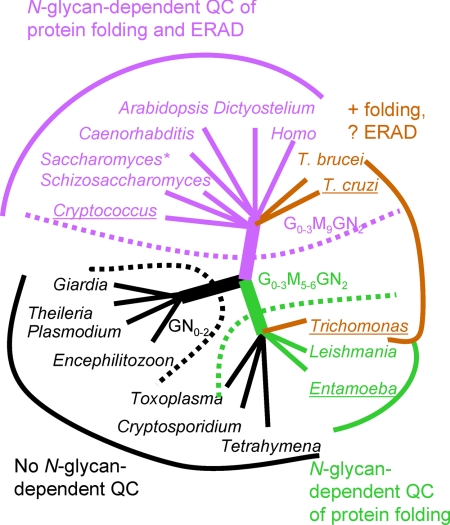
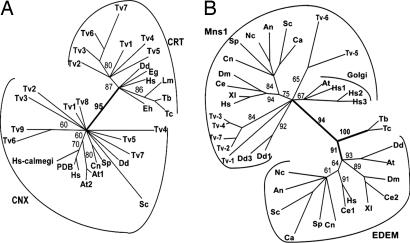
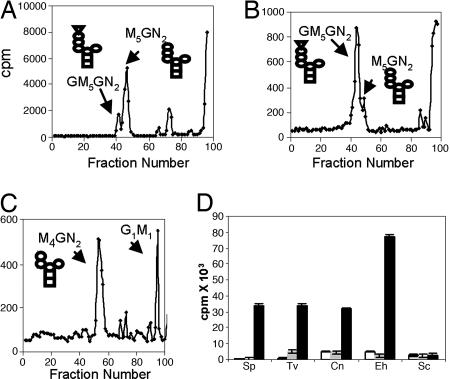
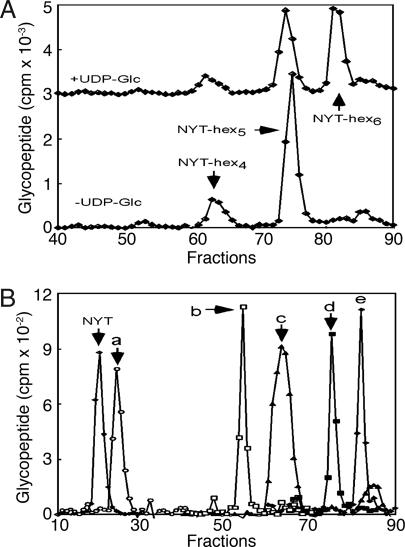
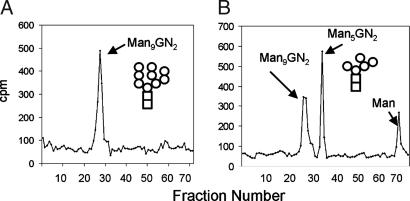
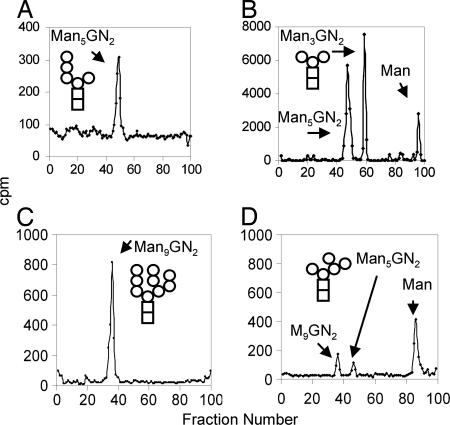
Asn-linked glycans (N-glycans) play important roles in the quality control (QC) of glycoprotein folding in the endoplasmic reticulum (ER) lumen and in ER-associated degradation (ERAD) of proteins by cytosolic proteasomes. A UDP-Glc:glycoprotein glucosyltransferase glucosylates N-glycans of misfolded proteins, which are then bound and refolded by calreticulin and/or calnexin in association with a protein disulfide isomerase. Alternatively, an alpha-1,2-mannosidase (Mns1) and mannosidase-like proteins (ER degradation-enhancing alpha-mannosidase-like proteins 1, 2, and 3) are part of a process that results in the dislocation of misfolded glycoproteins into the cytosol, where proteins are degraded in the proteasome. Recently we found that numerous protists and fungi contain 0-11 sugars in their N-glycan precursors versus 14 sugars in those of animals, plants, fungi, and Dictyostelium. Our goal here was to determine what effect N-glycan precursor diversity has on N-glycan-dependent QC systems of glycoprotein folding and ERAD. N-glycan-dependent QC of folding (UDP-Glc:glycoprotein glucosyltransferase, calreticulin, and/or calnexin) was present and active in some but not all protists containing at least five mannose residues in their N-glycans and was absent in protists lacking Man. In contrast, N-glycan-dependent ERAD appeared to be absent from the majority of protists. However, Trypanosoma and Trichomonas genomes predicted ER degradation-enhancing alpha-mannosidase-like protein and Mns1 orthologs, respectively, each of which had alpha-mannosidase activity in vitro. Phylogenetic analyses suggested that the diversity of N-glycan-dependent QC of glycoprotein folding (and possibly that of ERAD) was best explained by secondary loss. We conclude that N-glycan precursor length has profound effects on N-glycan-dependent QC of glycoprotein folding and ERAD.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
