

Genomics and the evolution, pathogenesis, and diagnosis of tuberculosis
- PMID: 17607348
- PMCID: PMC1904327
- DOI: 10.1172/JCI31810
Genomics and the evolution, pathogenesis, and diagnosis of tuberculosis
Abstract
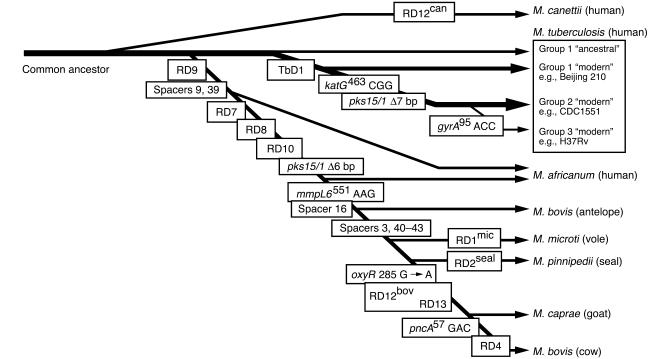
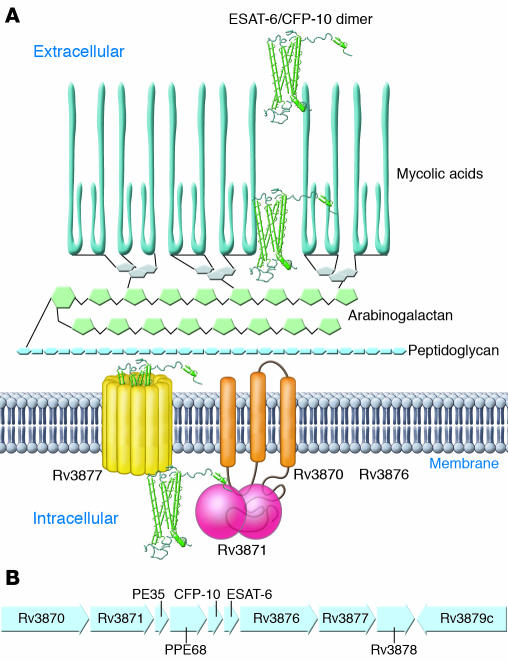
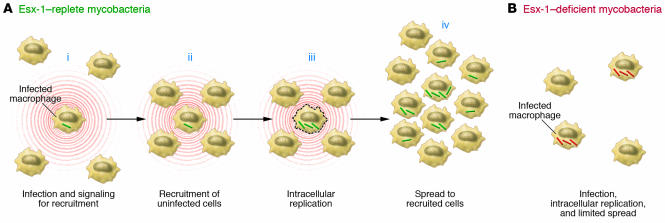
Tuberculosis kills nearly 2 million people annually, and current approaches to tuberculosis control are expensive, have limited efficacy, and are vulnerable to being overcome by extensively drug-resistant strains of Mycobacterium tuberculosis. Determination of the genome sequence of M. tuberculosis has revolutionized tuberculosis research, contributed to major advances in the understanding of the evolution and pathogenesis of M. tuberculosis, and facilitated development of new diagnostic tests with increased specificity for tuberculosis. In this review, we describe some of the major progress in tuberculosis research that has resulted from knowledge of the genome sequence and note some of the problems that remain unsolved.
Figures
References
-
- Smith N.H., et al. Ecotypes of the Mycobacterium tuberculosis complex. J. Theor. Biol. 2006;239:220–225. - PubMed
