

Topological reorganization of odor representations in the olfactory bulb
- PMID: 17608564
- PMCID: PMC1904499
- DOI: 10.1371/journal.pbio.0050178
Topological reorganization of odor representations in the olfactory bulb
Abstract
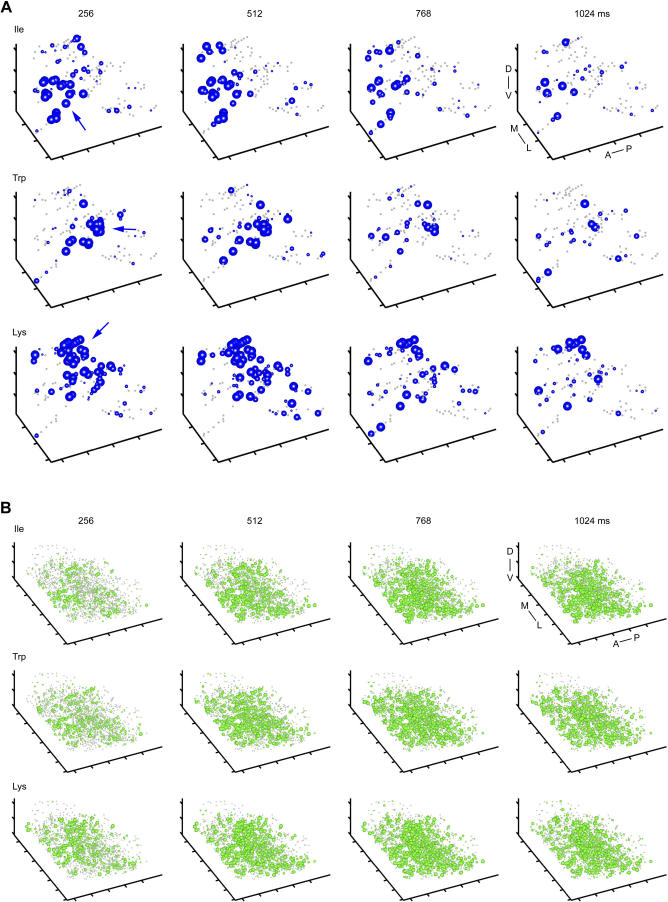
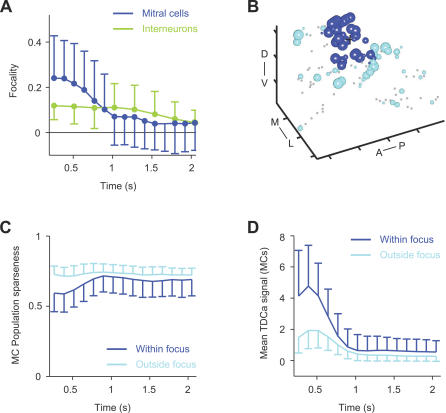
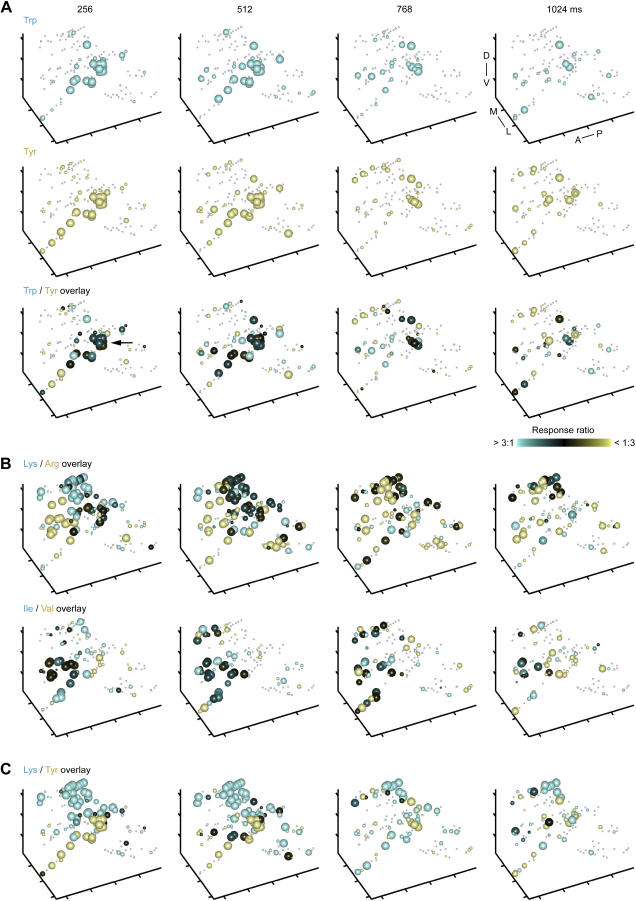
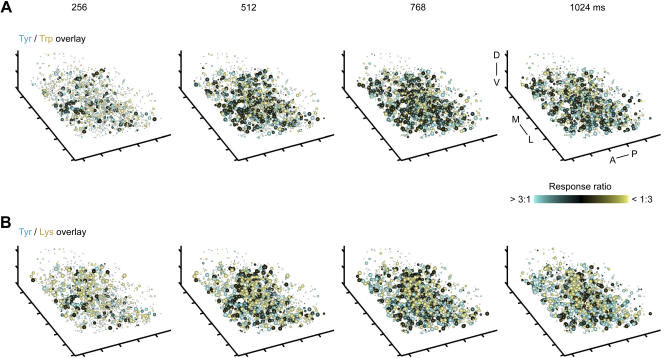
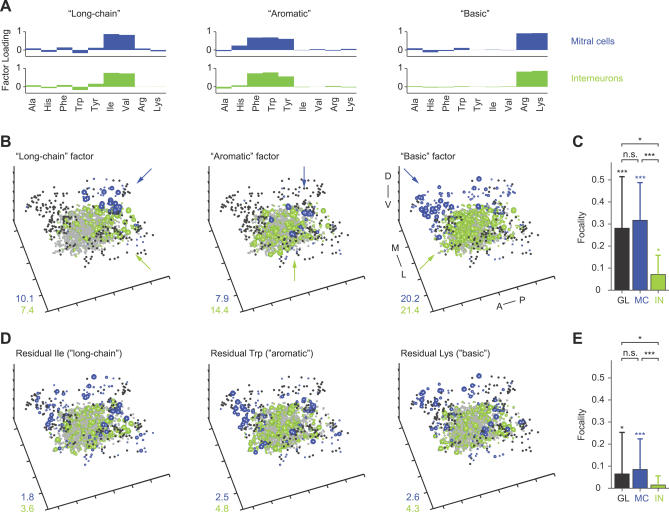
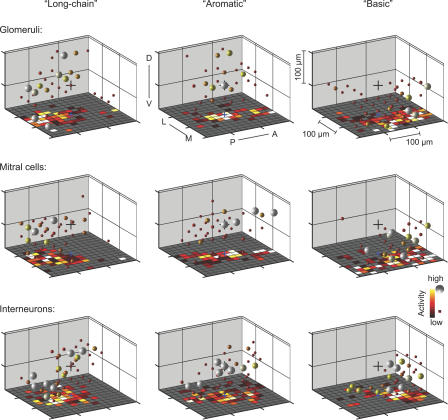
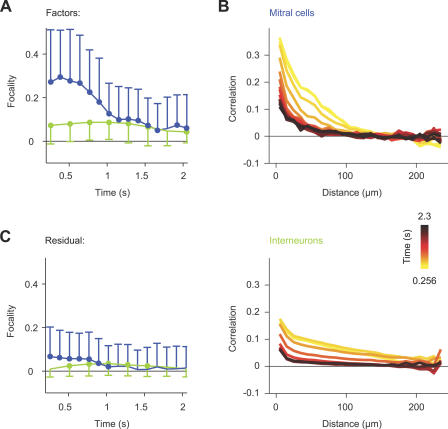
Odors are initially represented in the olfactory bulb (OB) by patterns of sensory input across the array of glomeruli. Although activated glomeruli are often widely distributed, glomeruli responding to stimuli sharing molecular features tend to be loosely clustered and thus establish a fractured chemotopic map. Neuronal circuits in the OB transform glomerular patterns of sensory input into spatiotemporal patterns of output activity and thereby extract information about a stimulus. It is, however, unknown whether the chemotopic spatial organization of glomerular inputs is maintained during these computations. To explore this issue, we measured spatiotemporal patterns of odor-evoked activity across thousands of individual neurons in the zebrafish OB by temporally deconvolved two-photon Ca(2+) imaging. Mitral cells and interneurons were distinguished by transgenic markers and exhibited different response selectivities. Shortly after response onset, activity patterns exhibited foci of activity associated with certain chemical features throughout all layers. During the subsequent few hundred milliseconds, however, MC activity was locally sparsened within the initial foci in an odor-specific manner. As a consequence, chemotopic maps disappeared and activity patterns became more informative about precise odor identity. Hence, chemotopic maps of glomerular input activity are initially transmitted to OB outputs, but not maintained during pattern processing. Nevertheless, transient chemotopic maps may support neuronal computations by establishing important synaptic interactions within the circuit. These results provide insights into the functional topology of neural activity patterns and its potential role in circuit function.
Conflict of interest statement
Figures
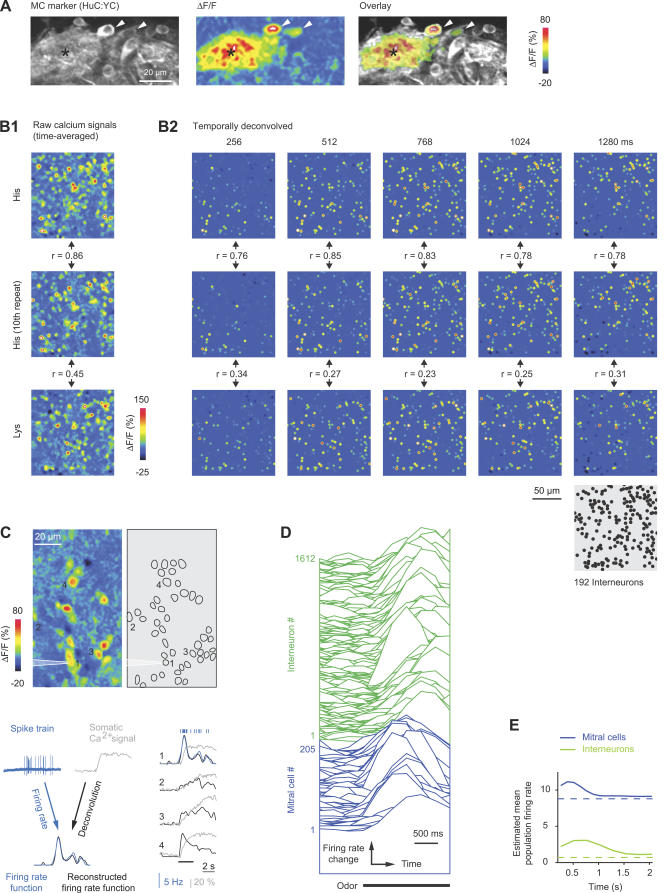
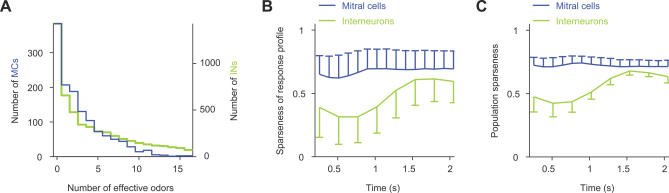
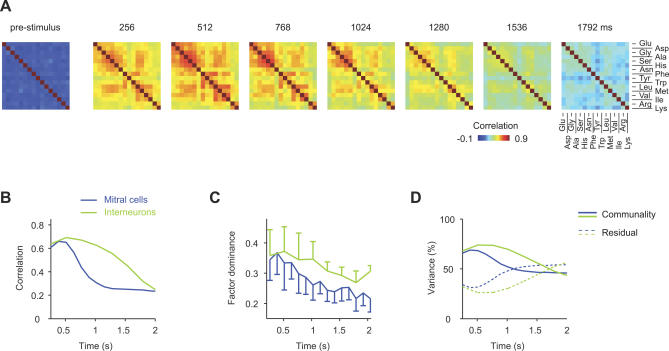
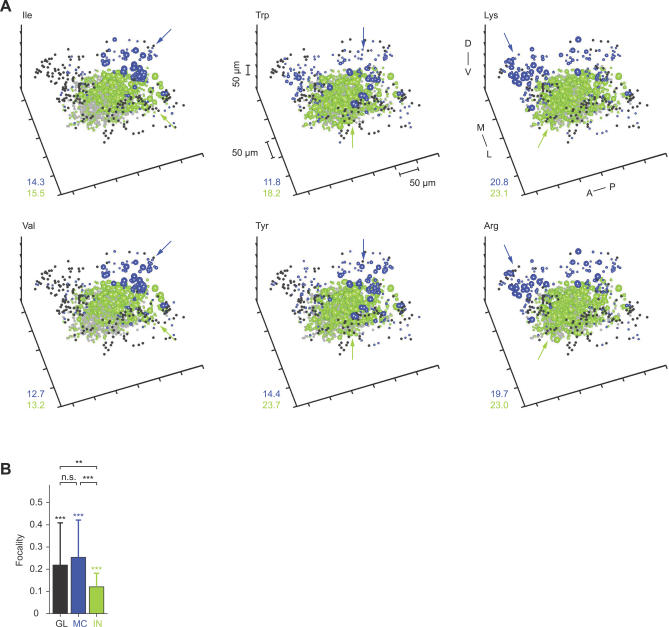
References
-
- Van Essen DC, Gallant JL. Neural mechanisms of form and motion processing in the primate visual system. Neuron. 1994;13:1–10. - PubMed
-
- Konishi M, Takahashi TT, Wagner H, Sullivan WE, Carr CE. Neurophysiological and anatomical substrates of sound localization in the owl. In: Edelman GM, Gall WE, Cowan WM, editors. Auditory function: neurobiological bases of hearing. New York: Wiley; 1988. pp. 721–745.
-
- Kaas JH. Topographic maps are fundamental to sensory processing. Brain Res Bull. 1997;44:107–112. - PubMed
-
- Wilson RI, Mainen ZF. Early events in olfactory processing. Annu Rev Neurosci. 2006;29:163–201. - PubMed
-
- Araneda RC, Kini AD, Firestein S. The molecular receptive range of an odorant receptor. Nature Neurosci. 2000;3:1248–1255. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
