

Productive human immunodeficiency virus type 1 infection in peripheral blood predominantly takes place in CD4/CD8 double-negative T lymphocytes
- PMID: 17609262
- PMCID: PMC2045436
- DOI: 10.1128/JVI.00492-07
Productive human immunodeficiency virus type 1 infection in peripheral blood predominantly takes place in CD4/CD8 double-negative T lymphocytes
Abstract
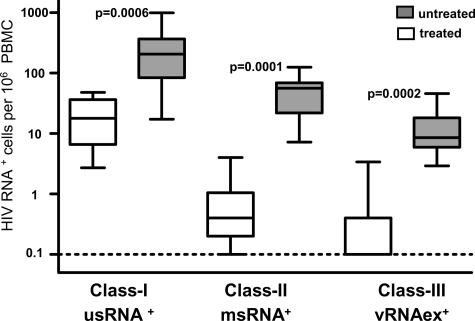
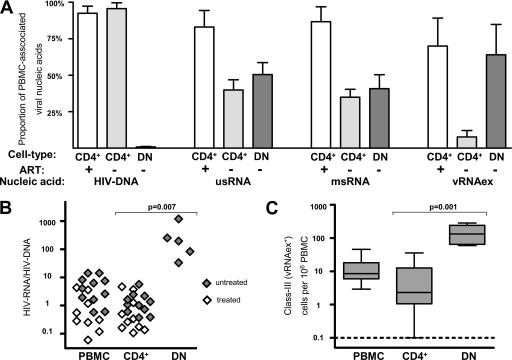
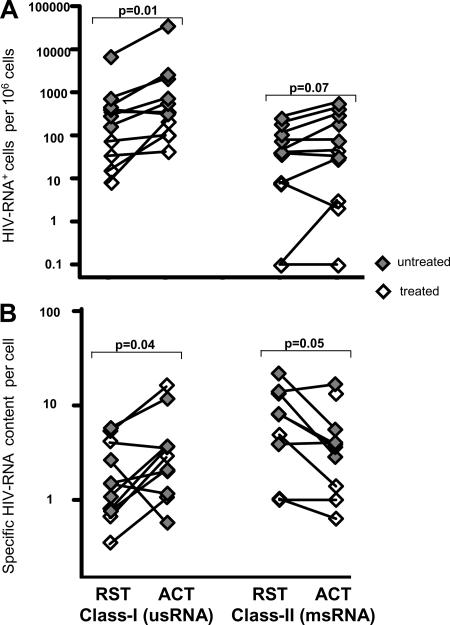
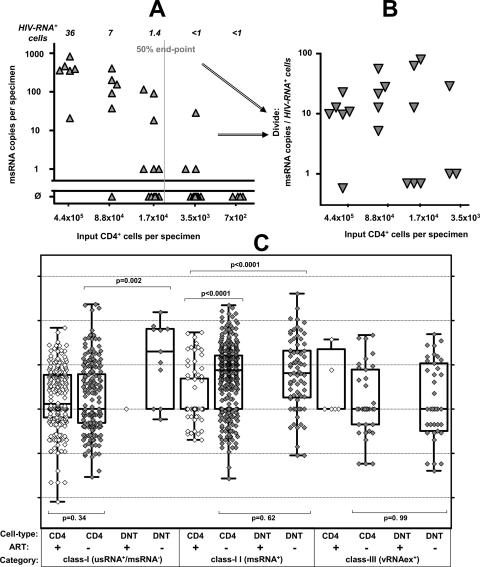
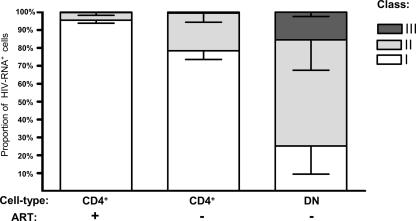
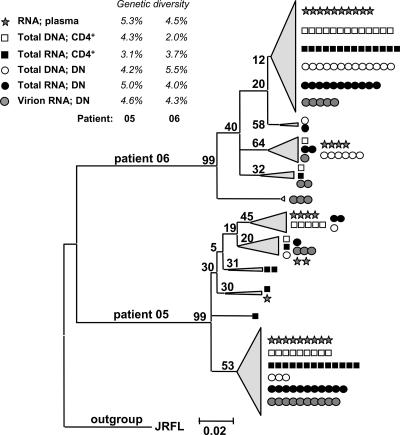
Human immunodeficiency virus type 1 (HIV-1) transcription is subject to substantial fluctuation during the viral life cycle. Due to the low frequencies of HIV-1-infected cells, and because latently and productively infected cells collocate in vivo, little quantitative knowledge has been attained about the range of in vivo HIV-1 transcription in peripheral blood mononuclear cells (PBMC). By combining cell sorting, terminal dilution of intact cells, and highly sensitive, patient-specific PCR assays, we divided PBMC obtained from HIV-1-infected patients according to their degree of viral transcription activity and their cellular phenotype. Regardless of a patient's treatment status, the bulk of infected cells exhibited a CD4+ phenotype but transcribed HIV-1 provirus at low levels, presumably insufficient for virion production. Furthermore, the expression of activation markers on the surface of these CD4+ T lymphocytes showed little or no association with enhancement of viral transcription. In contrast, HIV-infected T lymphocytes of a CD4-/CD8- phenotype, occurring exclusively in untreated patients, exhibited elevated viral transcription rates. This cell type harbored a substantial proportion of all HIV RNA+ cells and intracellular viral RNAs and the majority of cell-associated virus particles. In conjunction with the observation that the HIV quasispecies in CD4+ and CD4-)/CD8- T cells were phylogenetically closely related, these findings provide evidence that CD4 expression is downmodulated during the transition to productive infection in vivo. The abundance of viral RNA in CD4-/CD8- T cells from viremic patients and the almost complete absence of viral DNA and RNA in this cell type during antiretroviral treatment identify HIV+ CD4-/CD8 T cells as the major cell type harboring productive infection in peripheral blood.
Figures
References
-
- Aiken, C., J. Konner, N. R. Landau, M. E. Lenburg, and D. Trono. 1994. Nef induces CD4 endocytosis: requirement for a critical dileucine motif in the membrane-proximal CD4 cytoplasmic domain. Cell 76:853-864. - PubMed
-
- Bailey, J. R., A. R. Sedaghat, T. Kieffer, T. Brennan, P. K. Lee, M. Wind-Rotolo, C. M. Haggerty, A. R. Kamireddi, Y. Liu, J. Lee, D. Persaud, J. E. Gallant, J. Cofrancesco, Jr., T. C. Quinn, C. O. Wilke, S. C. Ray, J. D. Siliciano, R. E. Nettles, and R. F. Siliciano. 2006. Residual human immunodeficiency virus type 1 viremia in some patients on antiretroviral therapy is dominated by a small number of invariant clones rarely found in circulating CD4+ T cells. J. Virol. 80:6441-6457. - PMC - PubMed
-
- Bisset, L. R., S. Bosbach, Z. Tomasik, H. Lutz, J. Schüpbach, and J. Böni. 2001. Quantification of in vitro retroviral replication using a one-tube real-time RT-PCR system incorporating direct RNA preparation. J. Virol. Methods 91:149-155. - PubMed
-
- Blankson, J. N., D. Persaud, and R. F. Siliciano. 2002. The challenge of viral reservoirs in HIV-1 infection. Annu. Rev. Med. 53:557-593. - PubMed
-
- Brenchley, J. M., B. J. Hill, D. R. Ambrozak, D. A. Price, F. J. Guenaga, J. P. Casazza, J. Kuruppu, J. Yazdani, S. A. Migueles, M. Connors, M. Roederer, D. C. Douek, and R. A. Koup. 2004. T-cell subsets that harbor human immunodeficiency virus (HIV) in vivo: implications for HIV pathogenesis. J. Virol. 78:1160-1168. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
