

Extended exposure to Sonic hedgehog is required for patterning the posterior digits of the vertebrate limb
- PMID: 17610861
- PMCID: PMC2100419
- DOI: 10.1016/j.ydbio.2007.05.030
Extended exposure to Sonic hedgehog is required for patterning the posterior digits of the vertebrate limb
Abstract
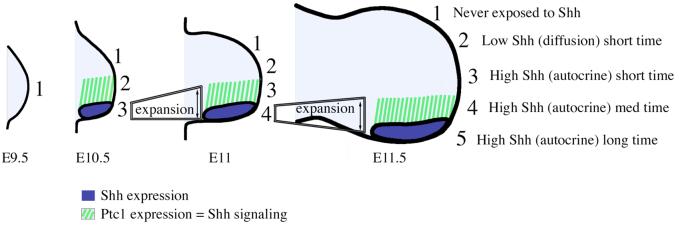
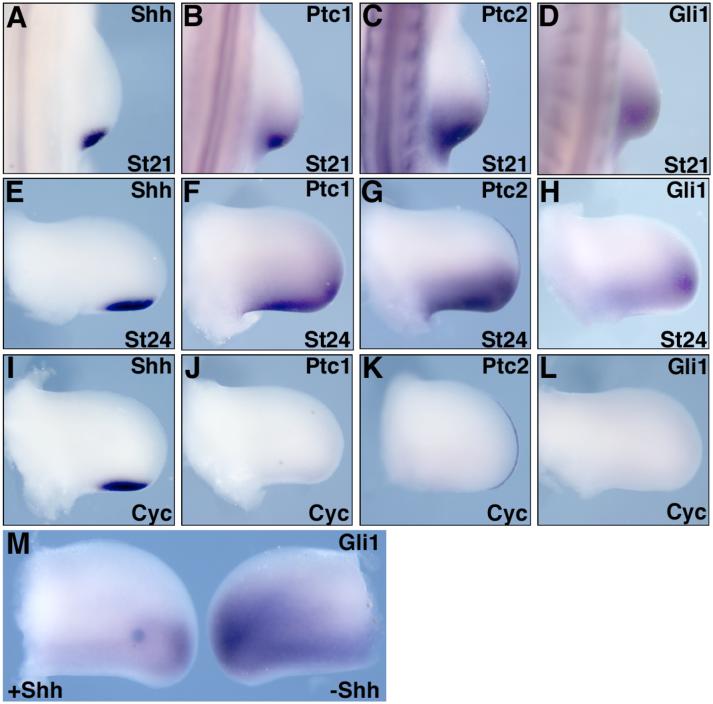
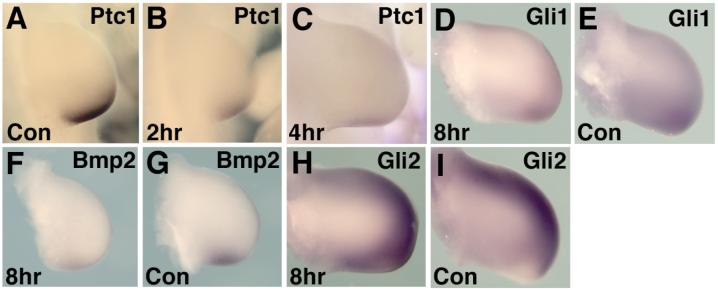
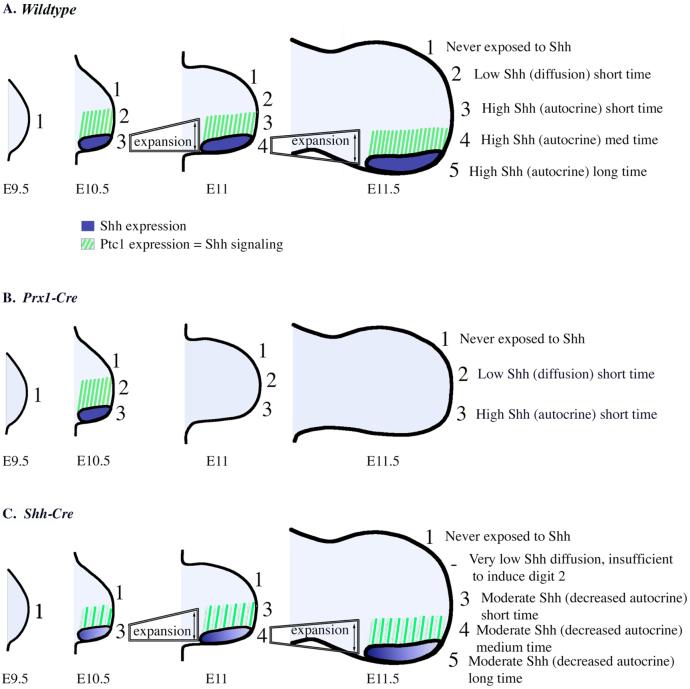
Sonic hedgehog (Shh) is a key signal in establishing different digit fates along the anterior-posterior axis of the vertebrate limb bud. Although the anterior digits appear to be specified by differential concentrations of Shh in a traditional, morphogen-like response, recent studies have suggested that posterior digits are specified by an extended time of exposure to Shh rather than, or in addition to, a threshold concentration of Shh. This model for digit patterning depends upon continued Shh signaling in the posterior limb through mid-to-late bud stages. We find that cyclopamine, a potent antagonist of Shh signaling, can down-regulate hedgehog target genes in the posterior limb throughout the time Shh is expressed, indicating that continued active Shh signaling indeed takes place. To further explore the relative roles of time and concentration of Shh during limb development, we carried out two additional series of experiments. To test the effect of limiting the time, but not the amount of Shh produced, we treated chick embryos with the hedgehog antagonist cyclopamine at various stages of limb development. We find that short exposures to Shh result in specification of only the most anterior digits and that more posterior digits are specified sequentially with increasing times of uninterrupted Shh activity. To test the effect of limiting the level of Shh produced, but not the time of exposure, we genetically modified Shh production in mice. As previously shown, reducing both the concentration of Shh produced and the duration of Shh exposure results in a loss of posterior digits. We find that maintaining a low level of Shh production throughout the normal time frame of ZPA signaling results in a near complete restoration of the posterior-most digits. These data are consistent with, and lend additional support to, the model that concentration of Shh seen and duration of exposure both contribute to the dose-dependent specification of digit identities, but for the posterior-most digits the temporal component is the more critical parameter.
Figures



References
-
- Ahn S, Joyner AL. Dynamic changes in the response of cells to positive hedgehog signaling during mouse limb patterning. Cell. 2004;118:505–516. - PubMed
-
- Alberch P, Gale EA. Size dependence during the development of the amphibian foot. Colchicine-induced digital loss and reduction. J Embryol Exp Morphol. 1983;76:177–197. - PubMed
-
- Aza-Blanc P, Lin HY, Ruiz I Altaba A, Kornberg TB. Expression of the vertebrate Gli proteins in Drosophila reveals a distribution of activator and repressor activities. Development. 2000;127:4293–4301. - PubMed
-
- Bai CB, Stephen D, Joyner AL. All mouse ventral spinal cord patterning by hedgehog is Gli dependent and involves an activator function of Gli3. Dev Cell. 2004;6:103–115. - PubMed
-
- Chiang C, Litingtung Y, Harris MP, Simandl BK, Li Y, Beachy PA, Fallon JF. Manifestation of the limb prepattern: development in the absence of sonic hedgehog function. Dev Biol. 2001;236:421–435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
