

Structures of phi29 DNA polymerase complexed with substrate: the mechanism of translocation in B-family polymerases
- PMID: 17611604
- PMCID: PMC1933411
- DOI: 10.1038/sj.emboj.7601780
Structures of phi29 DNA polymerase complexed with substrate: the mechanism of translocation in B-family polymerases
Abstract
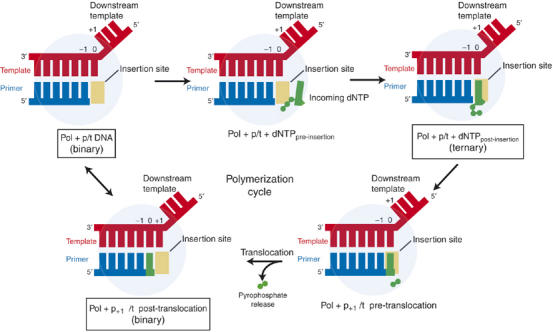
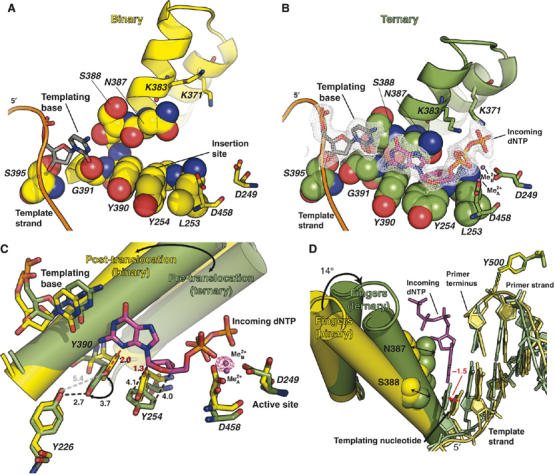
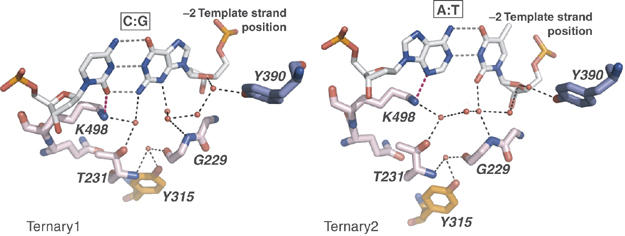
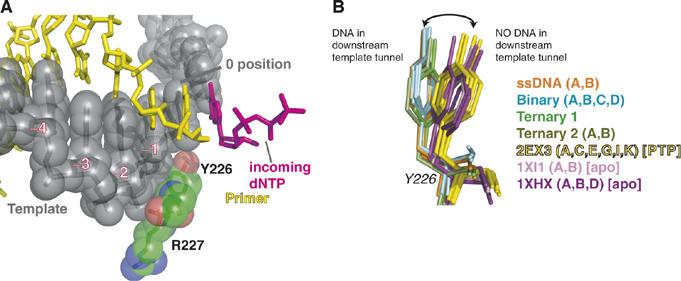
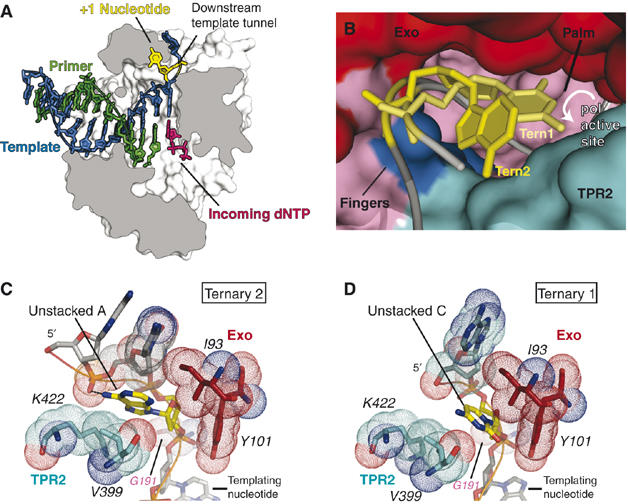
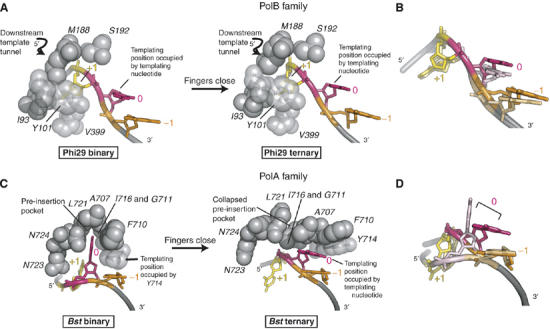
Replicative DNA polymerases (DNAPs) move along template DNA in a processive manner. The structural basis of the mechanism of translocation has been better studied in the A-family of polymerases than in the B-family of replicative polymerases. To address this issue, we have determined the X-ray crystal structures of phi29 DNAP, a member of the protein-primed subgroup of the B-family of polymerases, complexed with primer-template DNA in the presence or absence of the incoming nucleoside triphosphate, the pre- and post-translocated states, respectively. Comparison of these structures reveals a mechanism of translocation that appears to be facilitated by the coordinated movement of two conserved tyrosine residues into the insertion site. This differs from the mechanism employed by the A-family polymerases, in which a conserved tyrosine moves into the templating and insertion sites during the translocation step. Polymerases from the two families also interact with downstream single-stranded template DNA in very different ways.
Figures
References
-
- Beese LS, Friedman JM, Steitz TA (1993) Crystal structures of the Klenow fragment of DNA polymerase I complexed with deoxynucleoside triphosphate and pyrophosphate. Biochemistry 32: 14095–14101 - PubMed
-
- Blanco L, Salas M (1996) Relating structure to function in phi29 DNA polymerase. J Biol Chem 271: 8509–8512 - PubMed
-
- Blasco MA, Lázaro JM, Bernad A, Blanco L, Salas M (1992) Phi 29 DNA polymerase active site. Mutants in conserved residues Tyr254 and Tyr390 are affected in dNTP binding. J Biol Chem 267: 19427–19434 - PubMed
-
- Blasco MA, Lázaro JM, Blanco L, Salas M (1993) Phi 29 DNA polymerase active site. The conserved amino acid motif ‘Kx3NSxYG' is involved in template-primer binding and dNTP selection. J Biol Chem 268: 16763–16770 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
