

Differential susceptibility of yeast S and M phase CDK complexes to inhibitory tyrosine phosphorylation
- PMID: 17614281
- PMCID: PMC2034293
- DOI: 10.1016/j.cub.2007.05.075
Differential susceptibility of yeast S and M phase CDK complexes to inhibitory tyrosine phosphorylation
Abstract
Background: Several checkpoint pathways employ Wee1-mediated inhibitory tyrosine phosphorylation of cyclin-dependent kinases (CDKs) to restrain cell-cycle progression. Whereas in vertebrates this strategy can delay both DNA replication and mitosis, in yeast cells only mitosis is delayed. This is particularly surprising because yeasts, unlike vertebrates, employ a single family of cyclins (B type) and the same CDK to promote both S phase and mitosis. The G2-specific arrest could be explained in two fundamentally different ways: tyrosine phosphorylation of cyclin/CDK complexes could leave sufficient residual activity to promote S phase, or S phase-promoting cyclin/CDK complexes could somehow be protected from checkpoint-induced tyrosine phosphorylation.
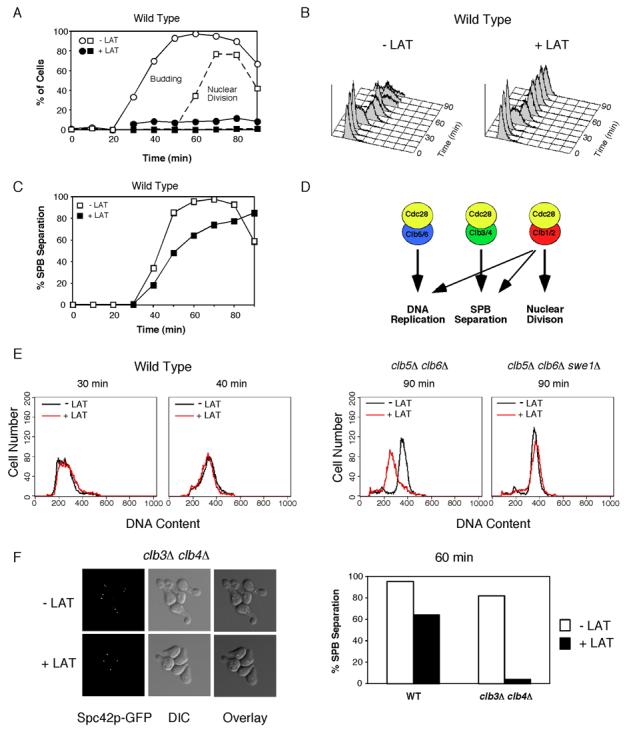
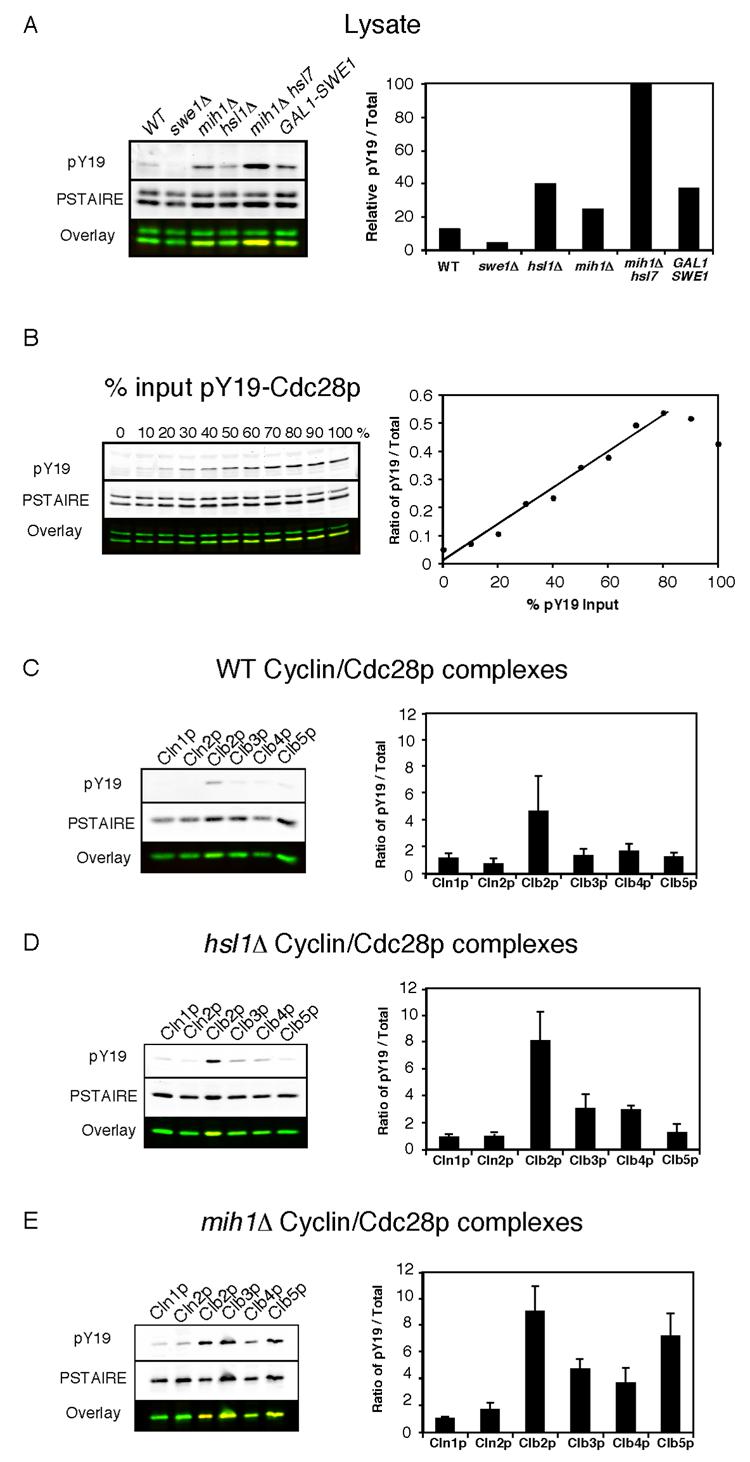
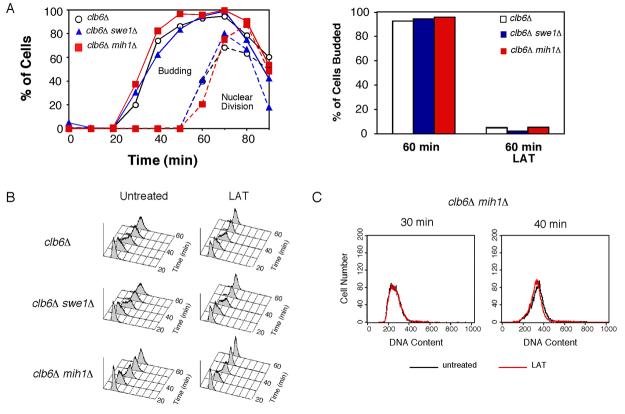
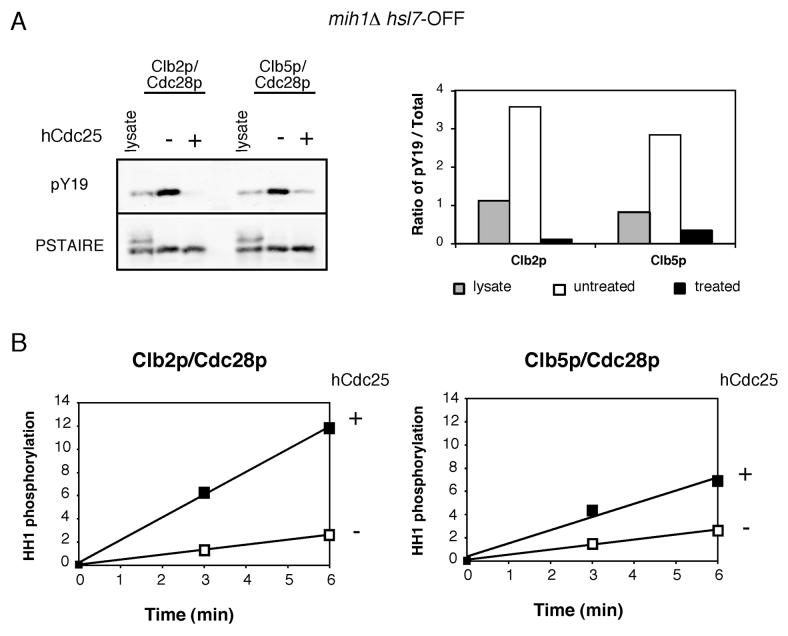
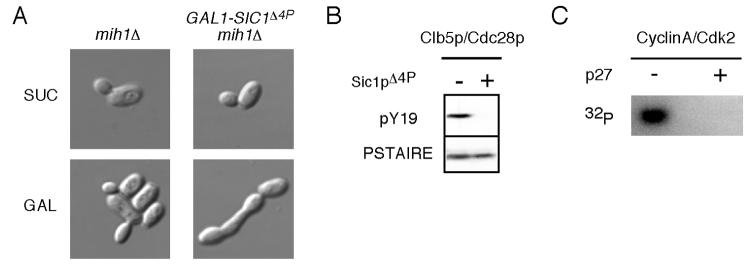
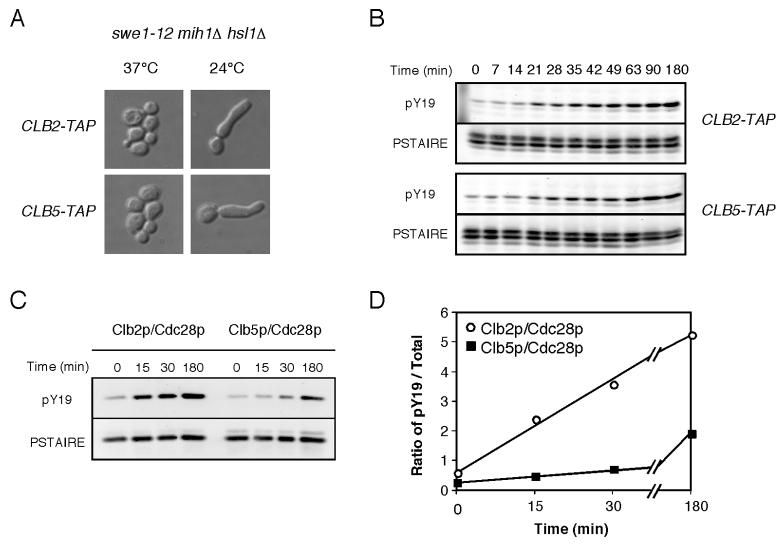
Results: We demonstrate that in Saccharomyces cerevisiae, several cyclin/CDK complexes are protected from inhibitory tyrosine phosphorylation, allowing Clb5,6p to promote DNA replication and Clb3,4p to promote spindle assembly, even under checkpoint-inducing conditions that block nuclear division. In vivo, S phase-promoting Clb5p/Cdc28p complexes were phosphorylated more slowly and dephosphorylated more effectively than were mitosis-promoting Clb2p/Cdc28p complexes. Moreover, we show that the CDK inhibitor (CKI) Sic1p protects bound Clb5p/Cdc28p complexes from tyrosine phosphorylation, allowing the accumulation of unphosphorylated complexes that are unleashed when Sic1p is degraded to promote S phase. The vertebrate CKI p27(Kip1) similarly protects Cyclin A/Cdk2 complexes from Wee1, suggesting that the antagonism between CKIs and Wee1 is evolutionarily conserved.
Conclusions: In yeast cells, the combination of CKI binding and preferential phosphorylation/dephosphorylation of different B cyclin/CDK complexes renders S phase progression immune from checkpoints acting via CDK tyrosine phosphorylation.
Figures
References
-
- Morgan DO. Principles of CDK regulation. Nature. 1995;374:131–134. - PubMed
-
- Humphrey T. DNA damage and cell cycle control in Schizosaccharomyces pombe. Mutat. Res. 2000;451:211–226. - PubMed
-
- Lew DJ. The morphogenesis checkpoint: how yeast cells watch their figures. Curr. Opin. Cell Biol. 2003;15:648–653. - PubMed
-
- Falck J, Mailand N, Syljuasen RG, Bartek J, Lukas J. The ATM-Chk2-Cdc25A checkpoint pathway guards against radioresistant DNA synthesis. Nature. 2001;410:842–847. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
