

Roles of phospholipase Cbeta and NMDA receptor in activity-dependent endocannabinoid release
- PMID: 17615097
- PMCID: PMC2277143
- DOI: 10.1113/jphysiol.2007.137497
Roles of phospholipase Cbeta and NMDA receptor in activity-dependent endocannabinoid release
Abstract
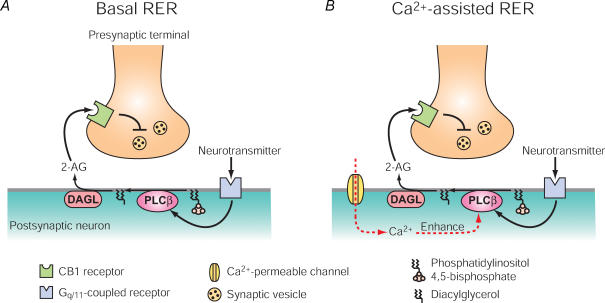
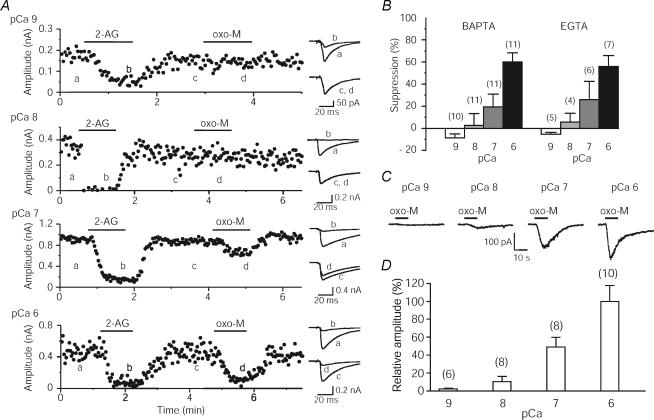
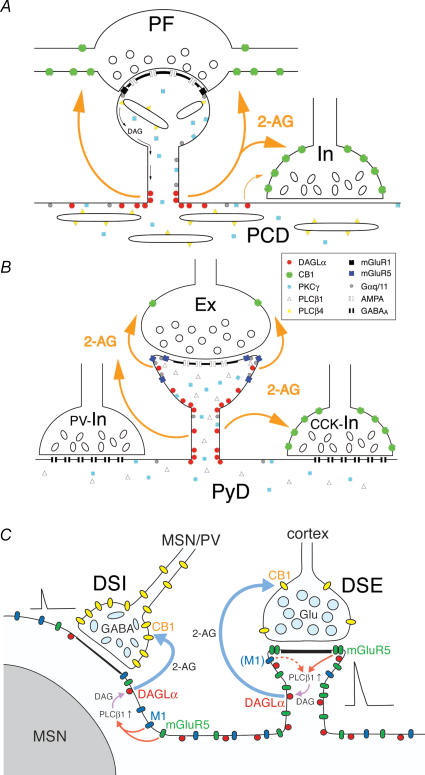
Endocannabinoids are released from postsynaptic neurons, activate presynaptic cannabinoid receptors and cause various forms of short-term and long-term synaptic plasticity throughout the brain. Using hippocampal and cerebellar neurons, we have revealed that endocannabinoid release can be induced through two different pathways. One is independent of phospholipase Cbeta (PLCbeta) and driven by Ca(2+) elevation alone (Ca(2+)-driven endocannabinoid release, CaER), and the other is PLCbeta-dependent and driven by activation of G(q/11)-coupled receptors (receptor-driven endocannabinoid release, RER). CaER is induced by activation of either voltage-gated Ca(2+) channels or NMDA receptors. RER is functional even at resting Ca(2+) levels (basal RER), but markedly enhanced by a small Ca(2+) elevation (Ca(2+)-assisted RER). In Ca(2+)-assisted RER, PLCbeta serves as a coincidence detector of receptor activation and Ca(2+) elevation. We have also demonstrated that Ca(2+)-assisted RER is essential for the endocannabinoid release triggered by synaptic activity. Our anatomical data show that a set of receptors and enzymes required for RER are well organized so that the excitatory input can trigger RER effectively. Certain forms of spike-timing-dependent plasticity (STDP) are reported to depend on endocannabinoid signalling. The NMDA receptor and PLCbeta might play key roles in the endocannabinoid-dependent forms of STDP as coincidence detectors with different timing dependences.
Figures
Similar articles
-
Ca(2+)-assisted receptor-driven endocannabinoid release: mechanisms that associate presynaptic and postsynaptic activities.Curr Opin Neurobiol. 2007 Jun;17(3):360-5. doi: 10.1016/j.conb.2007.03.012. Epub 2007 Apr 6. Curr Opin Neurobiol. 2007. PMID: 17419048 Review.
-
Phospholipase Cbeta serves as a coincidence detector through its Ca2+ dependency for triggering retrograde endocannabinoid signal.Neuron. 2005 Jan 20;45(2):257-68. doi: 10.1016/j.neuron.2005.01.004. Neuron. 2005. PMID: 15664177
-
Endocannabinoid signalling triggered by NMDA receptor-mediated calcium entry into rat hippocampal neurons.J Physiol. 2007 Oct 15;584(Pt 2):407-18. doi: 10.1113/jphysiol.2007.137505. Epub 2007 Jul 5. J Physiol. 2007. PMID: 17615096 Free PMC article.
-
Calcium signaling and synaptic modulation: regulation of endocannabinoid-mediated synaptic modulation by calcium.Cell Calcium. 2005 Sep-Oct;38(3-4):369-74. doi: 10.1016/j.ceca.2005.06.014. Cell Calcium. 2005. PMID: 16085309 Review.
-
Calcium influx through N-methyl-D-aspartate receptors triggers GABA release at interneuron-Purkinje cell synapse in rat cerebellum.Neuroscience. 2008 Jan 24;151(2):403-9. doi: 10.1016/j.neuroscience.2007.10.024. Epub 2007 Nov 4. Neuroscience. 2008. PMID: 18055124
Cited by
-
Dual mechanism of TRKB activation by anandamide through CB1 and TRPV1 receptors.PeerJ. 2019 Feb 21;7:e6493. doi: 10.7717/peerj.6493. eCollection 2019. PeerJ. 2019. PMID: 30809460 Free PMC article.
-
Cannabinoid-mediated short-term plasticity in hippocampus.J Comput Neurosci. 2014 Dec;37(3):533-47. doi: 10.1007/s10827-014-0518-4. Epub 2014 Aug 16. J Comput Neurosci. 2014. PMID: 25123173
-
Gene expression analysis indicates reduced memory and cognitive functions in the hippocampus and increase in synaptic reorganization in the frontal cortex 3 weeks after MDMA administration in Dark Agouti rats.BMC Genomics. 2018 Aug 2;19(1):580. doi: 10.1186/s12864-018-4929-x. BMC Genomics. 2018. PMID: 30071829 Free PMC article.
-
A biophysical model of endocannabinoid-mediated short term depression in hippocampal inhibition.PLoS One. 2013;8(3):e58926. doi: 10.1371/journal.pone.0058926. Epub 2013 Mar 18. PLoS One. 2013. PMID: 23527052 Free PMC article.
-
Electrophysiology of Endocannabinoid Signaling.Methods Mol Biol. 2023;2576:461-475. doi: 10.1007/978-1-0716-2728-0_38. Methods Mol Biol. 2023. PMID: 36152210
References
-
- Baude A, Nusser Z, Roberts JD, Mulvihill E, McIlhinney RA, Somogyi P. The metabotropic glutamate receptor (mGluR1α) is concentrated at perisynaptic membrane of neuronal subpopulations as detected by immunogold reaction. Neuron. 1993;11:771–787. - PubMed
-
- Brenowitz SD, Regehr WG. Associative short-term synaptic plasticity mediated by endocannabinoids. Neuron. 2005;45:419–431. - PubMed
-
- Brown SP, Brenowitz SD, Regehr WG. Brief presynaptic bursts evoke synapse-specific retrograde inhibition mediated by endogenous cannabinoids. Nat Neurosci. 2003;6:1048–1057. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous